

Caractéristiques des pores nucléaires, fonctions, composants
- 4216
- 1334
- Raphaël Meyer
Il pore nucléaire (Du grec, pores = étape ou transit) est la "porte" du noyau qui permet la majeure partie du transport entre la nucléoplasma et le cytoplasme. Le pore nucléaire rejoint les membranes internes et externes du noyau pour former des canaux, qui servent au transport des protéines et de l'ARN.
Le mot poro ne reflète pas la complexité de la structure en question. Pour cette raison, il est préférable de se référer au complexe de pores nucléaires (CPN), au lieu des pores nucléaires. Le CPN peut subir des changements dans sa structure pendant le transport ou les états du cycle cellulaire.
Récemment, il a été découvert que les nucléoporines, les protéines qui composent le CPN, ont un rôle important dans la régulation de l'expression des gènes. Ainsi, lorsque des mutations se produisent qui affectent la fonction des nucléoporines, des pathologies sont produites chez l'homme, comme l'auto-immune, les cardiomyopathies, les infections virales et le cancer.
[TOC]
Caractéristiques
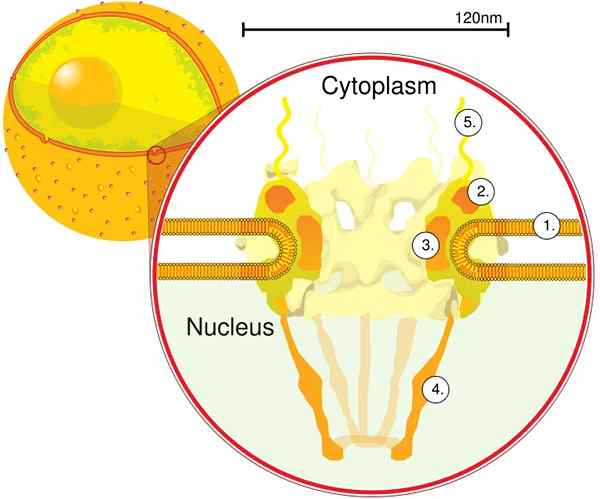
Grâce à la tomographie électronique, il a été déterminé que le CPN a une épaisseur de 50 nm, un diamètre externe entre 80 et 120 nm et un diamètre interne de 40 nm. De grandes molécules telles que la grande sous-unité de ribosomes (PM 1590 kDa) peuvent être exportées à l'extérieur du noyau par le CPN. On estime qu'il y a entre 2000 et 4000 cpn par noyau.
Le poids moléculaire d'un CPN individuel est d'environ 120 et 125 MDA (1 MDA = 106 Da) dans les vertébrés. En revanche, le CPN est plus petit dans les levures, dans lesquelles il a environ 60 MDA. Malgré la taille énorme du CPN, les nucléoporines sont très préservées dans tous les eucaryotes.
La translocalisation via le CPN est un processus rapide, dont la vitesse est de 1000 translocations / seconde. Cependant, le CPN ne détermine pas la direction du flux de transport lui-même.
Cela dépend du gradient Rangtp, qui est plus grand dans le noyau que dans le cytosol. Ce gradient est maintenu par un facteur d'échangeur de guanine Ran.
Pendant le cycle cellulaire, les CPN connaissent des cycles d'assemblage et de désunion. L'assemblage se produit à l'interface et immédiatement après la mitose.
Les fonctions
L'acide ribonucléique (petit ARN nucléaire, ARN messager, ARN de transfert), les protéines et les ribonucléoprotéines (RNP) doivent être activement transportés par CPN. Cela signifie que l'énergie de l'hydrolyse ATP et GTP est requise. Chaque molécule est transportée de manière spécifique.
En général, les molécules d'ARN sont remplies de complexes RNP formant des protéines, exportés de cette manière. En revanche, les protéines activement transportées vers le noyau doivent avoir un signe de séquence de localisation dans le noyau (SLN), possesseur de déchets d'acides aminés avec une charge positive (par exemple, KKKRK).
Peut vous servir: mésosomeLes protéines exportées vers le noyau doivent avoir un signal d'exportation vers le noyau (NES) riche en acide aminé de leucine.
En plus de faciliter le transport entre le noyau et le cytoplasme, les CPN sont impliqués dans l'organisation de la chromatine, la régulation de l'expression des gènes et la réparation de l'ADN. Les nucléoporines (NUP) favorisent l'activation ou la répression de la transcription, quel que soit l'état de prolifération cellulaire.
Dans les levures, les nupts se trouvent dans le CNP de l'emballage nucléaire. Dans les métazoaires, ils sont à l'intérieur. Ils remplissent les mêmes fonctions dans tous les eucaryotes.
Importation de substances
Grâce au CPN, il y a une diffusion passive de petites molécules dans les deux directions et le transport actif, l'importation de protéines, l'exportation d'ARN et de ribonucléoprotéines (RNP) et la navette des molécules bidirectionnelles. Ce dernier comprend l'ARN, le RNP et les protéines impliquées dans la signalisation, la biogenèse et le remplacement.
L'importation de protéines dans le noyau se produit en deux étapes: 1) la liaison des protéines au côté cytoplasmique du CPN; 2) Translocalisation dépendante de l'ATP à travers le CPN. Ce processus nécessite une hydrolyse ATP et l'échange de GTP / PIB entre le noyau et le cytoplasme.
Selon un modèle de transport, le complexe de protéines de réception se déplace le long du canal à travers l'Union, la dissociation et l'union aux séquences FG répétées des nucléoporines. De cette façon, le complexe passe d'une nucléoporine à une autre à l'intérieur du CPN.
Exportation de substances
Est similaire à l'importation. Ran GTPase impose la directionnalité au transport via CNP. Ran est un commutateur moléculaire avec deux états conformationnels, selon qu'il est lié au PIB ou au GTP.
Deux protéines régulatrices spécifiques déclenchent la conversion entre les deux états: 1) la protéine d'activation de la GTPase cytosolique (GAP), qui produit une hydrolyse GTP et convertit ainsi RAN-GTP en RAN-GDP; et 2) Facteur d'échange de guanina nucléaire (GEF), qui favorise l'échange du PIB par GTP et convertit Ran GDP en RAN-GTP.
Le cytosol contient principalement RAN GDP. Le noyau contient principalement RAN-GTP. Ce gradient des deux formes de conformation de RAN dirige le transport dans l'adresse appropriée.
L'importation du récepteur, avec la position, est facilitée par le syndicat aux répétitions-FG. Si vous atteignez le côté nucléaire du CNP, Ran-GTP rejoint le récepteur pour libérer votre position. Ainsi, Ran-GTP crée la direction du processus d'importation.
L'exportation nucléaire est similaire. Cependant, Ran-GTP dans le noyau favorise l'union de la position au récepteur d'exportation. Lorsque le récepteur d'exportation se déplace à travers le pore vers le cytosol, il rencontre Ran-Gap, ce qui induit une hydrolyse GTP au PIB. Enfin, le récepteur est libéré de sa position et Ran GDP dans le cytosol.
Peut vous servir: leucocytes polymorphonucléairesTransport de l'ARN
L'exportation de certains types d'ARN est similaire à l'exportation des protéines. Par exemple, ARNT et RNAN (petit nucléaire) utilisent le gradient RangTP et sont transportés par le CPN par des moyens. L'exportation des ribosomes matures dépend également du gradient RangTP.
L'ARNm est exporté d'une manière très différente des protéines et d'autres ARN. Pour son exportation, l'ARNm forme un complexe RNP Messenger (RNPM), dans lequel une molécule RNM est entourée de centaines de molécules de protéines. Ces protéines ont le traitement, Plafonnement, Empalme et polyadéntilly de MNA.
La cellule doit être capable de faire la distinction entre RNPM avec RNM et RNPm matures avec l'ARNm immature. L'ARNm, qui forme le complexe RPNM, pourrait adopter des topologies qui doivent être rénovées pour le transport. Avant que le RNPM entre dans le CPN, une étape de contrôle réalisée par des complexes de protéines de tramp et d'exosome se produit.
Lorsque la RNPM mature est assemblée, le RPNM est transporté par le canal au moyen d'un récepteur de transport (NXF1-NXT1). Ce récepteur a besoin d'hydrolyse ATP (pas un gradient RangTP) pour établir le remodelage de la directionnalité du RNPM, qui atteindra le cytoplasme.
Le complexe de pores nucléaires et le contrôle de l'expression des gènes
Certaines études indiquent que les composants CPN pourraient influencer la régulation de l'expression génétique par le contrôle de la structure de la chromatine et son accessibilité aux facteurs de transcription.
Dans les eucaryotes d'une évolution plus récente, l'hétérochromatine est préférentiellement située à la périphérie du noyau. Ce territoire est interrompu par les canaux euchromatines, qui sont maintenus par le panier nucléaire CPN. L'association du panier nucléaire avec l'euchromatine est liée à la transcription des gènes.
Par exemple, l'activation de la transcription au niveau du CPN implique l'interaction des composants du panier nucléaire avec des protéines telles que la saga d'histone acétyltransférase et les facteurs d'exportation d'ARN.
Ainsi, le panier nucléaire est une plate-forme pour de nombreux gènes de maintenance (Entretien ménager) hautement transcrit et gènes fortement induits par les changements dans les conditions environnementales.
Le complexe de pores nucléaires et de virologie
L'infection virale sur les cellules eucaryotes dépend du CPN. Dans chaque cas d'infection virale, son succès dépend de l'ADN, de l'ARN ou du RPN passant par le CPN pour atteindre son objectif ultime, qui est la réplication du virus.
Peut vous servir: inclusions cytoplasmiques: quelles sont les caractéristiques, les fonctionsLe virus 40 (SV40) a été l'un des modèles les plus étudiés pour étudier la fonction du CPN dans la transloalisation dans le noyau. C'est parce que SV40 a un petit génome (5.000 bases).
Il a été démontré que le transport de l'ADN virus est facilité les protéines protéiques du virus, qui protègent le virus jusqu'à ce que le noyau atteigne.
Composants
Le CPN est intégré à l'intérieur de l'enveloppe nucléaire et est composé entre environ 500 et 1000 nupts. Ces protéines sont organisées en sous-complexes ou modules structurels, qui interagissent les uns avec les autres.
Le premier module est un composant central, ou anneau, à l'intérieur du pore en forme d'une horloge de sable, qui est limité par un autre anneau de 120 nm de diamètre des deux côtés, intranucléaire et cytoplasmique. Le deuxième module est le noyau et les anneaux de cytoplasme (120 nm de diamètre chacun) situés autour du composant en forme de Sandwate.
Le troisième module comprend huit filaments qui sont projetés à partir de l'anneau de 120 nm dans la nucléoplasma et forment une structure en forme de panier. Le quatrième module est composé des filaments qui sont projetés sur le côté du cytoplasme.
Le complexe en forme Y, composé de six NUP et de protéines SEH 1 et Sec 13, est le complexe le plus grand et le mieux caractérisé du CNP. Ce complexe est l'unité essentielle qui fait partie de l'échafaudage CPN.
Malgré la faible similitude entre les séquences NUPS, l'échafaudage CPN est très préservé dans tous les eucaryotes.
Les références
- Beck, m., Blessé, e. 2016. Le complexe de pores nucléaires: comprendre sa fonction à travers une perspicacité structurelle. Revues de la nature, biologie des cellules moléculaires, doi: 10.1038 / nrm.2016.147.
- Ibarra, un., Hetzer, m.W. 2015. Protéines des pores nucléaires et contrôle des fonctions du génome. Gènes et développement, 29, 337-349.
- Kabachinski, G., Schwartz, t.OU. 2015. Le complexe de pores nucléaires - structure et fonction en un coup d'œil. Journal of Cell Science, 128, 423-429.
- Knocknhauer, k.ET., Schwartz, t.OU. 2016. Le complexe de pores nucléaires comme porte flexible et dynamique. Cell, 164, 1162-1171.
- Ptak, c., Aitchison, J.D., Wozniak, R.W. 2014. Le nucléaire multifonctionnel par complexe: une plate-forme pour contrôler l'expression des gènes. Opinion actuelle de la biologie cellulaire, DOI: 10.1016 / j.CEB.2014.02.001.
- Stawicki, S.P., Steffen, J. 2017. Républication: le complexe de pores nucléaires - une revue complète de la structure et de la fonction. Journal international de médecine académique, 3, 51-59.
- Tran, E.J., Went, S.R. 2006. Nuclear dynamique par complexes: la vie sur le bord. Cell, 125, 1041-1053.
- « Caractéristiques, structure, fonctions des smozymes
- Cycle cardiaque de phase et ses caractéristiques »