

Stades de phosphorylation oxydative, fonctions et inhibiteurs
- 2196
- 290
- Lucas Schneider
La la phosphorylation oxydative C'est un processus où les molécules ATP sont synthétisées à partir de l'ADP et du PToi (Phosphate inorganique). Ce mécanisme est réalisé par des bactéries et des cellules eucaryotes. Dans les cellules eucaryotes, la phosphorylation est réalisée dans la matrice mitochondriale des cellules non photosynthétiques.
La production d'ATP est dirigée par le transfert d'électrons des coenzymes NADH ou Fadh2 aussi2. Ce processus représente la plus grande production d'énergie dans la cellule et est dérivée de la dégradation des glucides et des graisses.
Source: robot8a [cc by-sa 4.0 (https: // CreativeCommons.Org / licences / by-sa / 4.0)] L'énergie stockée dans la charge et les gradients de pH, également connus sous le nom de force motrice protonique, permet de réaliser ce processus. Le gradient de protons généré, ce qui fait que la partie externe de la membrane a une charge positive en raison de la concentration de protons (H+) et la matrice mitochondriale est négative.
[TOC]
Où la phosphorylation oxydative se produit?
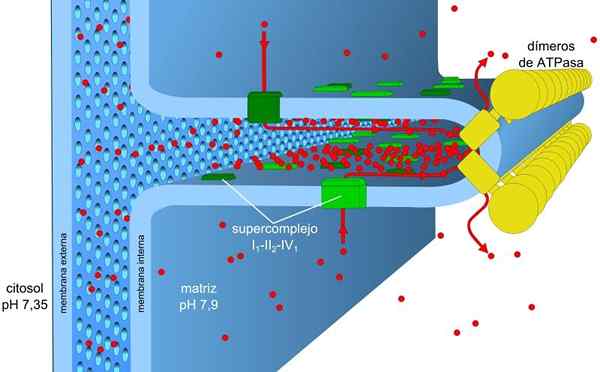
Les processus de transport d'électrons et de phosphorylation oxydative sont associés à une membrane. Dans les procaryotes, ces mécanismes sont effectués à travers la membrane plasmique. Dans les cellules eucaryotes, elles sont associées à la membrane des mitochondries.
Le nombre de mitochondries trouvées dans les cellules varie selon le type de cellule. Par exemple, chez les mammifères, les érythrocytes n'ont pas ces organites, tandis que d'autres types de cellules, tels que les cellules musculaires, peuvent en avoir des millions.
La membrane mitochondriale se compose d'une simple membrane externe, d'une membrane interne un peu plus complexe, et au milieu de l'espace intermembranal, où de nombreuses enzymes dépendant de l'ATP sont situées.
La membrane externe contient une protéine appelée Porine qui forme les canaux pour la diffusion simple de petites molécules. Cette membrane est responsable du maintien de la structure et de la forme des mitochondries.
La membrane interne a une plus grande densité et est riche en protéines. Il est également étanche pour les molécules et les ions, donc, pour le traverser, ils ont besoin de protéines intermembranales qui les transportent.
Dans la matrice, les plis de la membrane interne s'étendent, formant des crêtes qui lui permettent d'avoir une grande surface dans un petit volume.
Central Central
Les mitochondries sont considérées comme le centre d'énergie cellulaire. Il y a les enzymes impliquées dans les processus du cycle d'acide cytrique, de l'oxydation des acides gras et des enzymes redox et des protéines du transport d'électrons et de la phosphorylation de PhD.
Le gradient de concentration des protons (gradient de pH) et le gradient de charges ou de potentiel électrique dans la membrane interne des mitochondries sont la cause de la force motrice prottonique. La petite perméabilité de la membrane interne pour les ions (autre que H+) permet aux mitochondries d'avoir un gradient de tension stable.
Transport électronique, pompage des protons et ATP obtenant simultanément dans les mitochondries, grâce à la force motrice protonique. Le gradient de pH maintient des conditions acides dans la matrice intermembranaire et mitochondriale avec des conditions alcalines.
Pour deux électrons transférés à l'O2 Environ 10 protons sont pompés par la membrane, créant un gradient électrochimique. L'énergie détachée dans ce processus est produite progressivement en passant des électrons à travers la chaîne du convoyeur.
Étapes
L'énergie libérée lors des réactions de réduction de l'oxyde de NADH et FADH2 Il est considérablement élevé (environ 53 kcal / mol pour chaque paire d'électrons), donc pour être utilisé dans la fabrication de molécules ATP, il doit être produit progressivement avec le passage d'électrons à travers des transporteurs.
Ceux-ci sont organisés dans quatre complexes situés dans la membrane mitochondriale interne. Le couplage de ces réactions à la synthèse d'ATP est effectué dans un cinquième complexe.
Peut vous servir: b lymphocytes: caractéristiques, structure, fonctions, typesChaîne de transport d'électrons
NADH transfère quelques électrons qui entrent dans le complexe I de la chaîne de transport d'électrons. Les électrons sont transférés au mononucléotide de flavine, puis à l'ubiquinone (Coenzyme Q) à travers un convoyeur en sucre en fer. Ce processus libère une grande quantité d'énergie (16,6 kcal / mol).
Ubiquinona transporte les électrons à travers la membrane vers le complexe III. Dans ce complexe, les électrons passent par les cytochromes B et C1 Merci à un transporteur de sucre de fer.
À partir du complexe III, les électrons passent au complexe IV (cytochrome c oxydase), transféré un par cytochrome c (protéine membranaire périphérique). Dans le complexe IV, les électrons passent à travers une paire d'ions de cuivre (Cupour2+), puis au cytochrome cpour, puis à une autre paire d'ions de cuivre (Cub2+) et de cela au cytochrome à3.
Enfin, les électrons sont transférés à o2 qui est le dernier accepteur et forme une molécule d'eau (h2O) Pour chaque paire d'électrons reçus. Le passage des électrons du complexe IV à O2 Il génère également beaucoup d'énergie libre (25,8 kcal / mol).
Succinato Coq réductase
Le complexe II (Coq réductase succinate) reçoit une paire d'électrons du cycle de l'acide citrique, en raison de l'oxydation d'une molécule de succinate à fumarate. Ces électrons sont transférés à la mode, puis en passant par un groupe de fer-azufre, à l'Ubiquinona. De cette coenzyme, ils vont au complexe III et suivent l'itinéraire décrit précédemment.
L'énergie libérée dans la réaction de transfert d'électrons à la mode ne suffit pas pour augmenter les protons à travers la membrane, de sorte que dans cette étape de la chaîne, il n'y a pas de force motrice prottonique, et par conséquent le Fadh donne moins de H+ que le nadh.
Couplage énergétique ou transduction
L'énergie générée dans le processus de transport d'électrons décrit précédemment, doit être utilisée pour la production d'ATP, la réaction catalysée par l'ATP synthase ou l'enzyme complexe V. La conservation d'une telle énergie est connue sous le nom de couplage d'énergie, et le mécanisme a été difficile à caractériser.
Plusieurs hypothèses ont été décrites pour décrire cette transduction d'énergie. Le plus accepté est l'hypothèse de couplage chimiosmotique, décrite ci-dessous.
Couplage chimiosmotique
Ce mécanisme propose que l'énergie utilisée pour la synthèse d'ATP provient d'un gradient protonique dans les membranes cellulaires. Ce processus intervient dans les mitochondries, les chloroplastes et les bactéries et est lié au transport d'électrons.
Les complexes I et IV du transport électronique agissent comme des pompes à protons. Ceux-ci souffrent de changements conformationnels qui leur permettent de pomper les protons vers l'espace intermembranal. Dans le complexe IV pour chaque paire d'électrons, deux protons sont pompés à l'extérieur de la membrane et deux autres restent dans la matrice formant H2SOIT.
Ubiquinona dans le complexe III accepte les protons des complexes I et II et les libère à l'extérieur de la membrane. Les complexes I et III permettent chacun le passage de quatre protons pour chaque paire d'électrons transportés.
La matrice mitochondriale a une faible concentration de protons et un potentiel électrique négatif tandis que l'espace intermembranal présente les conditions inverses. Le flux de protons à travers cette membrane implique le gradient électrochimique qui stocke l'énergie nécessaire (± 5 kcal / mol par proton) pour la synthèse d'ATP.
Synthèse ATP
L'enzyme ATP synthétase est le cinquième complexe impliqué dans la phosphorylation oxydative. Est responsable de profiter de l'énergie du gradient électrochimique pour former l'ATP.
Cette protéine transmembraale se compose de deux composants: f0 et f1. Le composant f0 permet le retour des protons à la matrice mitochondriale fonctionnant comme un canal et le F1 catalyse la synthèse de l'ATP via ADP et PToi, en utilisant l'énergie dudit retour.
Peut vous servir: protéines de convoyeur à membrane: fonctions et typesLe processus de synthèse de l'ATP nécessite un changement structurel en F1 et l'assemblage des composants F0 et f1. La translocation de protons à travers f0 Il provoque des changements conformationnels dans trois sous-unités de F1, permettant à cela d'agir comme un moteur de rotation, dirigeant la formation de l'ATP.
La sous-unité qui est responsable de l'union de l'ADP avec PToi passe d'un état faible (L) à un actif (t). Lorsque l'ATP est formé, une deuxième sous-unité passe à un état ouvert (O) qui permet la libération de cette molécule. Après la libération de l'ATP, cette sous-unité passe de l'état ouvert à un état inactif (L).
Les molécules ADP et PToi Ils rejoignent une sous-unité qui est passée d'un État ou de l'État L.
produits
La chaîne de convoyeurs d'électrons et la phosphorylation produisent des molécules d'ATP. L'oxydation du NADH produit environ 52,12 kcal / mol (218 kJ / mol) d'énergie libre.
La réaction globale de l'oxydation du NADH est:
NADH + 1⁄2 ou2 +H+ ↔ H2O + NAD+
Le transfert d'électrons de Nadh et Fadh2 Il se produit à travers plusieurs complexes, permettant au changement d'énergie libre ΔG ° d'être divisé en «packages» plus petits, qui sont couplés à la synthèse d'ATP.
L'oxydation d'une molécule NADH génère la synthèse de trois molécules d'ATP. Tandis que l'oxydation d'une molécule fadh2 Il est attaché à la synthèse de deux ATP.
Ces coenzymes proviennent des processus de glycolyse et de cycle d'acide citrique. Pour chaque molécule de glucose dégradée, 36 ou 38 molécules ATP se terminent, selon l'emplacement des cellules. Dans le cerveau et les muscles squelettiques, 36 ATP sont produits tandis que 38 ATP sont produits dans le tissu musculaire.
Les fonctions
Tous les organismes, unicellulaires et multicellulaires, ont besoin d'une énergie minimale dans leurs cellules pour effectuer les processus en leur sein, et à leur tour maintenir des fonctions vitales dans le corps complet.
Les processus métaboliques nécessitent une énergie pour être effectuée. La majeure partie de l'énergie utilisable est obtenue par la dégradation des glucides et des graisses. Cette énergie dérive du processus de phosphorylation oxydatif.
Contrôle de la phosphorylation oxydative
La vitesse d'utilisation de l'ATP dans les cellules, contrôle la synthèse de la même chose, et à son tour, en raison du couplage de la phosphorylation oxydative avec la chaîne de transport d'électrons, il régule généralement le taux de transport électronique.
La phosphorylation oxydative a un contrôle strict qui garantit que l'ATP n'est pas généré plus rapidement qu'il n'est consommé. Il existe certaines étapes dans le processus de transport d'électrons et la phosphorylation couplée qui régulent le taux de production d'énergie.
Contrôle coordonné de la production d'ATP
Les principales voies de production d'énergie (ATP cellulaire) sont la glycolyse, le cycle de l'acide citrique et de la phosphorylation oxydative. Le contrôle coordonné de ces trois processus régule la synthèse de l'ATP.
Le contrôle de la phosphorylation par le rapport d'action de masse ATP dépend de la contribution précise des électrons dans la chaîne de transport. Cela dépend à son tour de la relation [nadh] / [nad+qui est préservé élevé par l'action de la glycolyse et du cycle d'acide citrique.
Ce contrôle coordonné est effectué en régulant les points de contrôle de la glycolyse (le citrate inhibé par le PFK) et le cycle d'acide cytrique (pyruvate de déshydrogénase, citrate de coupe, itchitrate déshydrogénase et α-zétoglutarate déshydrogénase).
Contrôle de l'accepteur
Le complexe IV (cytochrome c oxydase) est une enzyme régulée par l'un de ses substrats, c'est-à-dire que son activité est contrôlée par le cytochrome C réduit (C2+), qui à son tour est en équilibre avec la relation des concentrations entre [NADH] / [NAD+] et le rapport d'action de masse de [ATP] / [ADP] + [PToi]].
Il peut vous servir: ton: caractéristiques et fonctionsPlus [nadh] / [nad+] et abaisser le [ATP] / [ADP] + [PToi], plus de concentration sera le cytochrome [c2+] et plus sera l'activité du complexe IV. Ceci est interprété, par exemple, si nous comparons les organismes avec différentes activités de repos et une activité élevée.
Chez un individu avec une activité physique élevée, la consommation d'ATP et donc son hydrolyse à ADP + PToi Il sera très élevé, générant une différence dans le rapport d'action de masse qui provoque une augmentation de [C2+et donc une augmentation de la synthèse de l'ATP. Dans un individu au repos, la situation inverse se produit.
En fin de compte, la vitesse de la phosphorylation oxydative augmente avec la concentration d'ADP dans les mitochondries. Cette concentration dépend des translocteurs ADP-ATP en charge du transport des nucléotides adénine et PToi Du cytosol à la matrice mitochondriale.
Agents décacoplants
La phosphorylation oxydative est influencée par certains agents chimiques, qui permettent au transport électronique de se poursuivre sans la phosphorylation de l'ADP, découplant la production et la conservation de l'énergie.
Ces agents stimulent la vitesse de consommation d'oxygène des mitochondries en l'absence d'ADP, provoquant également une augmentation de l'hydrolyse de l'ATP. Ils agissent en éliminant un intermédiaire ou en cassant un état d'énergie de la chaîne de convoyeurs d'électrons.
Le 2.4-dinitrofénol, un acide faible qui passe à travers les membranes mitochondriales, est responsable de la dissipation du gradient de protons, car ils les rejoignent du côté acide et les libèrent du côté de base.
Ce composé a été utilisé comme une "pilule minceur" car il a été constaté qu'il produisait une augmentation de la respiration, par conséquent, une augmentation du taux métabolique et une perte de poids associée. Cependant, il a été démontré que son effet négatif pouvait même provoquer la mort.
La dissipation du gradient protonique produit de la chaleur. Les cellules tissulaires adipeuses brunes utilisent le découplage, contrôlé hormonalement, pour produire de la chaleur. Les mammifères d'hibernate et les nouveau-nés qui manquent de cheveux sont constitués de ce tissu qui les sert comme une sorte de couverture thermique.
Inhibiteurs
Les composés ou agents inhibiteurs empêchent à la fois la consommation de ou2 (transport électronique) comme la phosphorylation oxydative associée. Ces agents évitent la formation d'ATP par l'utilisation d'énergie produite dans le transport électronique. Par conséquent, la chaîne de transport s'arrête car une telle consommation d'énergie n'est pas organisée.
L'antibiotique d'oligomicine fonctionne comme un inhibiteur de la phosphorylation dans de nombreuses bactéries, empêchant la stimulation de l'ADP de la synthèse de l'ATP.
Il y a aussi des agents ionophoreux, qui composent des complexes solubles en matières grasses avec des cations telles que k+ et na+, et passer à travers la membrane mitochondriale avec ces cations. Les mitochondries utilisent ensuite l'énergie produite dans le transport électronique pour pomper des cations au lieu de synthétiser l'ATP.
Les références
- Alberts, B., Bray, D., Hopkin, k., Johnson, un., Lewis, J., Raff, m., Roberts, k. & Walter, P. (2004). Biologie cellulaire essentielle. New York: Garland Science.
- Cooper, G. M., Hausman, R. ET. & Wright, n. (2010). La cellule. (PP. 397-402). Marbán.
- Devlin, t. M. (1992). Manuel de biochimie: avec des corrélations cliniques. John Wiley & Sons, Inc.
- Garrett, R. H., & Grisham, C. M. (2008). Biochimie. Thomson Brooks / Cole.
- Lodish, H., Darnell, J. ET., Berk, un., Kaiser, C. POUR., Krieger, m., Scott, M. P., & Matsudaira, P. (2008). Biologie cellulaire molleculaire. Macmillan.
- Nelson, D. L., & Cox, M. M. (2006). Principes de lehninger de la biochimie 4e édition. Ed Omega. Barcelone.
- Voet, D., & Voet, J. g. (2006). Biochimie. Élégant. Pan -American Medical.
- « Caractéristiques d'Euglenophyta, reproduction, nutrition, classification
- Formation, fonctions et types de glucosides »