

Structure d'uracile, fonctions, propriétés, synthèse
- 3092
- 517
- Adrien Remy
Il Uracile Il s'agit d'une base d'azote de pyrimidine, qui se trouve dans l'acide ribonucléique (ARN). C'est l'une des caractéristiques qui différencient l'ARN de l'acide désoxyribonucléique (ADN), car ce dernier a Timina au lieu de l'uracile. Les deux substances, l'uracile et le timina, ne diffèrent que que le second a un groupe méthyle.
Du point de vue évolutif, il a été proposé que l'ARN était la première molécule qui stockait des informations génétiques et travaillait comme catalyseur dans les cellules, avant l'ADN et les enzymes. Pour cette raison, on pense que l'uracile a eu un rôle clé dans l'évolution de la vie.
Source: Kemikungen [domaine public] Chez les êtres vivants, l'uracile n'est pas trouvé librement, mais généralement des nucléotides monophosphates (UMP), du diphosphate (UDP) et du trphosphate (UTP). Ces nucléotides d'uracile ont des fonctions différentes, telles que la biosynthèse de l'ARN et le glycogène, l'interconversion isomère des sucres et la régulation de la synthamine synthase.
[TOC]
Structure et propriétés
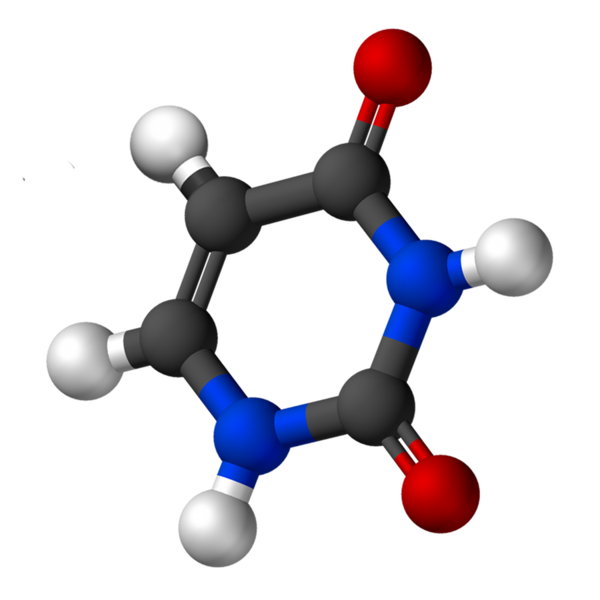
L'uracile, appelé 2.4-dioxipiridine, a la formule empirique C4H4N2SOIT2, dont le poids moléculaire est de 112,09 g / mol et est purifié comme une poudre blanche.
La structure de l'uridine est un anneau hétérocyclique avec quatre atomes de carbone et deux azotés, avec des liaisons doubles alternatives. C'est planaire.
Il a une solubilité de 50 mg / ml, 25 ºC, dans un hydroxyde de sodium 1 m et un PKA entre 7.9 et 8.2. La longueur d'onde où son absorbance maximale se produit (ʎMax) C'est entre 258 et 260 nm.
Biosynthèse
Il existe un chemin commun pour la biosynthèse des nucléotides de pyrimidine (uracile et cytokines). La première étape est la biosynthèse du phosphate de carbamoil de CO2 et NH4+, qui est catalysé par le phosphate de carbamoil synthétase.
La pyrimidine est construite à partir de phosphate de glucide et d'aspartate. Les deux substances réagissent et forment n-carbamoilaspartato, réaction catalysée par l'aspartate de Transcabamoilasa (atcasa) (atcasa). La fermeture de l'anneau de pyrimidine peut être déshydratée par la dihydrootase et produit du l-dihydrotatoate.
Le l-dihydrotoate est oxydé et transformé en orotate; L'accepteur d'électrons est le NAD+. C'est une réaction catalysée par la dihydroorotate déshydrogénase. L'étape suivante consiste dans le transfert du groupe Fosforibosyl, du phosphoribosyl pyrophosphate (PRPP) à orotate. La forme du gradilatato (WPO) et le piroffosphat inorganique (PPI), catalysé par la transfertas de fosforibosyl orotate.
La dernière étape consiste en une décarboxylation de l'anneau de pyrimidine du groupe (WPO). La forme uridilaato (uridine-5'-monophosphate, UMP), qui est catalysée par une décarboxylase.
Ensuite, par la participation d'une kinase, un groupe de phosphate est transféré de l'ATP à l'UMP, formant UDP (Uridine-5'-diphosphate). Ce dernier est répété, formant UTP (Uridine-5'-triffosphate).
Peut vous servir: branches de la biologie et que étudient-ilsRégulation de la biosynthèse
Dans les bactéries, la régulation de la biosynthèse de la pyrimidine se produit par une rétroaction négative, au niveau de l'aspartate Transcabamoilasa (ATCASA).
Cette enzyme est inhibée par le CTP (Citidin-5'-Trophosphate), qui est le produit final de la voie de biosynthèse des pyrimidines. ATCAS.
Chez les animaux, la régulation de la biosynthèse de la pyrimidine se produit par rétroaction négative, au niveau de deux enzymes: 1) le phosphate de carbamoil synthase II, qui est inhibé par UTP et activé par l'ATP et le PRPP; et 2) l'oMP discarboxylase, qui est inhibée par le produit de la réaction qu'elle catalyse elle-même, l'UMP. La vitesse de biosynthèse de l'OMP varie avec la disponibilité du PRPP.
Fonction dans la biosynthèse de l'ARN
L'uracile est présent dans tous les types d'ARN, comme l'ARN messager (MNA), l'ARN de transfert (ARNT) et l'ARN ribosomal (ARN). La biosynthèse de ces molécules se produit par un processus appelé transcription.
Pendant la transcription, les informations contenues dans l'ADN sont copiées dans l'ARN via un ARN PON polymérase. Le processus inverse, dans lequel les informations contenues dans l'ARN sont copiées dans l'ADN, se produit dans certains virus et plantes par la transcriptase inverse.
La biosynthèse de l'ARN nécessite des nucléosides de tryposphate (NTP), à savoir: Tryphosphate uridine (UTP), tryposphate cytidine (CTP), adénine tryposphate adénine (ATP) et tryphosphate guanine (GTP). La réaction est:
(ARN)n déchets + NTP -> (ARN)N + 1 déchets + PPI
L'hydrolyse du pyrophosphate inorganique (PPI) fournit de l'énergie pour la biosynthèse de l'ARN.
Fonction dans la biosynthèse du sucre
Les esters de sucre sont très courants dans les organismes vivants. Certains de ces esters sont les diphosphas des esters nucléosides, comme UDP-sucre, qui sont très abondants dans les cellules. L'UDP-sucre implique la biosynthèse des disaccharides, des oligosaccharides et des polysaccharides.
Dans les plantes, la biosynthèse du saccharose se produit de deux manières: un principal et un secondaire.
L'itinéraire principal consiste en le transfert du D-glucose de l'UDP-D-Glucosa à la D-Frocease pour former du saccharose et UDP. La route secondaire comprend deux étapes: il commence par l'UDP-D-glucose et le fructose-6-phosphate et culmine avec la formation de saccharose et de phosphate.
Dans les glandes mammaires, la biosynthèse du lactose se produit de l'UDP-D-galactose et du glucose.
Peut vous servir: branches de la biochimieChez les plantes, la biosynthèse de la cellulose est réalisée par condensation continue des déchets bêta-d-glycosyles, de l'UDP-glucose à l'extrémité non réductrice de la chaîne de polyglucose croissante. De même, la biosynthèse de l'amylose et de l'amylopectine nécessite un udp-glucose comme substrat donneur de glucose à la chaîne en croissance.
Chez les animaux, l'UDP-glucose et l'ADP-glucose sont utilisés pour la biosynthèse du glycogène. De même, la biosynthèse du sulfate de chondroïtine nécessite UDP-xylose, UDP-galactose et UDP-glucuronato.
Fonction dans l'interconversion isomère des sucres
La conversion du galactose en intermédiaire de glycolyse se produit à travers la route Leloir. L'une des étapes de cette route est catalysée par l'enzyme UDP-galactose-4-épimérase, qui facilite l'interconversion de l'UDP-galactose en UDP-glucose.
Fonction dans la biosynthèse des glycoprotéines
Pendant la biosynthèse des glycoprotéines, les protéines passent par les sacs CIS, moyens et trans de l'appareil Golgi.
Chacun de ces sacs a un ensemble d'enzymes qui traitent les glycoprotéines. Monomères de sucre, comme le glucose et le galactose.
Les nucléotides-hexose sont transportés dans les réservoirs de Golgi par Antiporte. L'UDP-galactose (UDP-Gal) et l'UDP-N-acétylgalactosamine (UDP-Galnac) entrent dans les réservoirs du cytosol par échange par UMP.
Dans le réservoir de Golgi, une phosphatase hydrolyse un groupe phosphate de la forme UDP et UMP et PI. L'UDP provient des réactions catalysées par la galactosyltransférase et la N-acétylgalactosamiltransférase. L'UMP formé par la phosphatase sert à échanger des nucléotides-hexose.
Fonction dans la régulation de la synthase
Un mécanisme de régulation de la glutamine synthase est la modification covalente, qui consiste en une adélilation, qui inactive et flanchée, qui l'a active. Cette modification covalente est réversible et catalysée par l'adényltransférase.
L'activité de l'adényltransférase est modulée par l'union de la protéine PII, qui est régulée par une modification covalente, l'uridinilation.
L'uridililation et le décalage sont effectués par l'uridililtransférase. Dans cette enzyme, l'activité d'uridillation est due à la glutamine et au phosphate, et est activée par l'union de l'alpha-zotoglutarate et de l'ATP à la PII.
Fonction dans l'édition ARN
Certains RNM sont modifiés avant la traduction. Dans certains organismes eucaryotes, comme Trucei trucei, Il y a l'édition de l'ARN de la transcription du gène de la sous-unité II de la cytochrome oxydase. Cela se produit en insérant les déchets d'uracile, une réaction catalysée par l'uridiltransférase terminale.
Peut vous servir: Curl: Composition, parties, importanceUn ARN guide, complémentaire au produit édité, agit comme tempéré pour le processus d'édition. Les paires de bases formées entre la transcription initiale et l'ARN guide impliquent des paires de bases g = u qui ne sont pas Watson-record et sont courantes dans l'ARN.
Biosynthèse UDP-glucose
Dans des conditions physiologiques, la biosynthèse du glycogène du glucose-1-phosphate est thermodynamiquement impossible (ΔG positif). Pour cette raison, avant la biosynthèse, l'activation du glucose-1-phosphate (G1P) se produit. Ce G1P et UTP combinent la réaction pour former du glucose diphosphate uridine (UDP-glucose ou UDPG).
La réaction est catalysée par la pyrophosphoryllase de l'UDP-glucose, et est la suivante:
G1p + UTP -> UDP -Glucosa + 2PI.
La variation énergétique libre de Gibbs dans cette étape est grande et négative (-33,5 kJ / mol). Pendant la réaction d'oxygène, le G1P attaque l'atome alpha-phosphore alforo de l'UDP-glucose et du pyrophosphate inorganique (PPI). Ensuite, l'IPP est hydrolysé par un pyrophosphates inorganiques, dont l'énergie d'hydrolyse est celle qui entraîne la réaction générale.
L'UDP-glucose est une substance "à haute énergie". Il permet de former les liaisons glycosidiques entre le résidu de glucose et la chaîne polysaccharide croissante. Ce même principe d'énergie est applicable aux réactions où participent le sucre UDP, comme la biosynthèse des disaccharides, des oligosaccharides et des glycoprotéines.
ADN uracile glycosilasa
Il y a des lésions d'ADN qui se produisent spontanément. L'une de ces lésions est le Sprontane. Dans ce cas, la réparation a lieu en raison de la base d'ADN modifiée au moyen d'une enzyme appelée ADN uracile glycosilasa.
L'enzyme de l'ADN de l'uracile glycosilasa élimine la cytokine endommagée (uracile), produisant un résidu désoxyribose qui n'a pas la base d'azote, appelée site AP (site apurinique-apirimidinique).
Ensuite, l'enzyme AP endonucléase fait une coupe dans le squelette phosphodiester du site AP, éliminant le résidu de sucre-phosphate. L'ADN polymérase I restaure le brin endommagé.
Les références
- Bohinski, R. 1991. Biochimie. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, t.M. 2000. Biochimie. Reverté éditorial, Barcelone.
- Lodish, H., Berk, un., Zipurski, S.L., Matsudaria, P., Baltimore, D., Darnell, J. 2003. Biologie cellulaire et moléculaire. Éditorial Medica Panamericana, Buenos Aires, Bogotá, Caracas, Madrid, Mexique, Sāo Paulo.
- Nelson, D. L., Cox, M. M. 2008. Lehninger-primeciples de biochimie. W.H. Freeman, New York.
- Voet, D. Et Voet, J. 2004. Biochimie. John Wiley and Sons, USA.
- « Formule de coût marginal, calcul et exercices
- Caractéristiques des pyrimidines, structure, fonctions »