

Caractéristiques des microfilaments, structure, fonctions, pathologie
- 4469
- 1187
- Noa Da silva
Les microfilaments o Les filaments d'actine, sont l'une des trois composantes fondamentales du cytosquelette des cellules eucaryotes.
Chez les eucaryotes, les gènes codant pour les microfilaments d'actine sont très conservés dans tous les organismes, ils sont donc souvent utilisés comme marqueurs moléculaires pour diverses études.
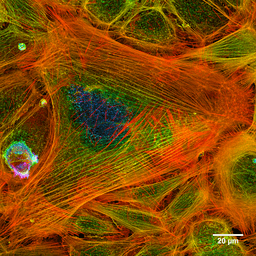
Photographie des filaments d'actine d'une cellule teinte (Source: Howard Vindin [CC BY-SA 4.0 (https: // CreativeCommons.Org / licences / by-sa / 4.0)] via Wikimedia Commons) Les microfilaments sont distribués dans tout le cytosol, mais ils sont particulièrement abondants dans la région sous-jacente de la membrane plasmique, où ils forment un réseau complexe et sont associés à d'autres protéines spéciales pour former le cytosquelette.
Les réseaux de microfilament dans le cytoplasme des cellules de mammifères sont codés par deux des six gènes décrits pour l'actine, qui sont impliqués dans la dynamique des microfilaments et qui sont même très importants lors de la différenciation des cellules souches.
De nombreux auteurs conviennent que les microfilaments sont les protéines les plus diverses, polyvalentes et importantes du cytosquelette de la plupart.
Dans ce type de cellules, cependant, il existe des filaments homologues en microfilaments, mais qui sont formés par une autre protéine: la protéine MREB.
À l'heure actuelle, on pense que le gène qui code pour cette protéine est un gène ancestral possible pour l'actine eucaryote. Cependant, l'homologie de la séquence d'acides aminés qui forme la protéine MREB n'est que de 15% par rapport à la séquence d'actine.
Tout comme un élément fondamental du cytosquelette, tout défaut phénotypique à la fois dans les microtubules et dans les filaments intermédiaires et les microfilaments d'actine (cytosquelette) peut provoquer différentes pathologies cellulaires et systémiques.
[TOC]
Caractéristiques et structure
Les microfilaments sont composés de monomères protéiques de la famille d'actine, qui sont des protéines contractiles extrêmement abondantes dans les cellules eucaryotes, car elles participent également à la contraction musculaire.
Ces filaments ont un diamètre entre 5 et 7 nm, ils sont donc également connus sous le nom de filaments minces et sont composés de deux formes d'actine: la forme globulaire (actine G) et la forme filamenteuse (actine f).
Les protéines qui participent au cytosquelette sont connues sous le nom d'actines γ et β, tandis que ceux qui participent à la contraction sont généralement des actines α.
Peut vous servir: cytoplasme: fonctions, parties et caractéristiquesLa proportion d'actine globulaire et d'actine filamenteuse dans le cytosol dépend des besoins cellulaires, car les microfilaments sont des structures très variables et polyvalentes, qui sont constamment de croissance et de raccourcissement en raison de la polymérisation et de la dépolymérisation.
L'actine G est une petite protéine globulaire, composée de près de 400 acides aminés et d'environ 43 kDa.
Les monomères d'actinement qui composent les microfilaments sont ordonnés sous la forme d'un volet hélicoïdal, car chacun souffre d'une torsion lorsqu'il est associé à ce qui suit.
L'actine G est associée à une molécule Ca2 + et à un autre ATP, qui stabilise sa forme globulaire; Tandis que l'actine F est obtenu après l'hydrolyse du phosphate terminal de la molécule d'ATP dans l'actine G, ce qui contribue à la polymérisation.
Organisation
Les filaments d'actine peuvent être organisés sous la forme de "poutres" ou de "réseaux" qui ont des fonctions différentes à l'intérieur des cellules. Les faisceaux forment des structures parallèles liées par des ponts croisés assez rigides.
Les réseaux, en revanche, sont des structures plus confortables, telles que des mailles à trois dimensions avec les propriétés des gels semi-solides.
Il existe de nombreuses protéines associées aux filaments d'actine ou de microfilament et qui sont appelés ABP (de l'anglais Protéines de liaison à l'actine), qui ont des sites spécifiques pour cela.
Beaucoup de ces protéines permettent aux microfilaments d'interagir avec les deux autres composants du cytosquelette: microtubules et filaments intermédiaires, ainsi qu'avec les autres composants de la face intérieure de la membrane plasmique.
Parmi les autres protéines interagissant avec les microfilaments, il y a des feuilles nucléaires et de la spectrine (dans les globules rouges).
Comment les filaments actin sont-ils formés?
Étant donné que les monomères globulaires de l'actine se joignent toujours à la même manière, orientés dans la même direction, les microfilaments ont une polarité définie, avec deux extrêmes: un "de plus" et un "moins".
La polarité de ces filaments est très importante, car elles se développent considérablement plus rapidement en raison de leur fin positive, où les nouveaux monomères actin g sont ajoutés.
 Représentation graphique des microfilaments d'actine (Source: Travail dérivé: Retama (talk) mince_filament_formation.SVG: Mikael Häggström [domaine public] via Wikimedia Commons)
Représentation graphique des microfilaments d'actine (Source: Travail dérivé: Retama (talk) mince_filament_formation.SVG: Mikael Häggström [domaine public] via Wikimedia Commons) La première chose qui se déroule lors de la polymérisation des filaments d'actine est un processus appelé «nucléation», qui se compose de l'association de trois monomères protéiques.
Peut vous servir: plasomolyseÀ cette coupe-tricot, de nouveaux monomères sont ajoutés par les deux extrémités, de sorte que le filament se développe. Les monomères d'actinent sont capables d'hydrolyz l'ATP à chaque union, ce qui a des implications dans la vitesse de la polymérisation, car les portions d'actine-ATP dissocient plus de difficulté que l'actine-ADP.
L'ATP n'est pas nécessaire pour la polymérisation et la fonction de béton de son hydrolyse n'a pas encore été clarifiée.
Certains auteurs considèrent que, puisque les événements de polymérisation de l'actine sont rapides.
Régulation
La polymérisation des filaments d'actine et sa dépolimérisation sont des processus hautement régulés par une série de protéines spécifiques, qui sont responsables du remodelage des filaments.
Exemple des protéines qui régulent les dépolymères. Une autre protéine, la prophylaïne, a une fonction opposée, car elle stimule l'association des monomères (en stimulant l'échange d'ADP pour l'ATP).
Les fonctions
Les microfilaments interagissent avec les filaments de myosine associés aux protéines transmembranaires qui ont un domaine dans le cytosol et une autre cellule à l'étranger, ils participent donc aux processus de mobilité cellulaire.
Ces microfilaments associés à la membrane plasmique médiatisent différentes réponses cellulaires à différents types de stimuli. Par exemple, l'adhésion cellulaire des tissus épithéliaux est dirigée par des protéines transmembranaires appelées cadherinas, qui interagissent avec les microfilaments pour recruter des facteurs de réponse.
Les filaments d'actine interagissent avec les filaments intermédiaires pour faire la transmission de stimuli extracellulaires à des endroits clés tels que les ribosomes et les chromosomes à l'intérieur du noyau.
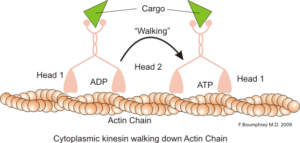 Représentation de la fonction motrice intracellulaire des microfilaments d'actine (Source: Boumphreyfr [CC BY-SA 3.0 (https: // CreativeCommons.Org / licences / by-sa / 3.0)] via Wikimedia Commons)
Représentation de la fonction motrice intracellulaire des microfilaments d'actine (Source: Boumphreyfr [CC BY-SA 3.0 (https: // CreativeCommons.Org / licences / by-sa / 3.0)] via Wikimedia Commons) Une fonction classique et très étudiée des microfilaments est sa capacité à former des «ponts», des «rails» ou des «autoroutes» pour le mouvement de la protéine Myosin Motor I, qui est capable de charger des vésicules de transport des organites vers le plasma membranaire sur la sécrétion de la sécrétion chemins.
Les microfilaments interagissent également avec la myosine II pour établir l'anneau contractile formé pendant la cytokinèse, précisément au cours de la dernière étape de la division cellulaire dans laquelle le cytosol est séparé de la tige et.
Peut vous servir: leucocytes polymorphonucléairesEn général, les microfilaments en forme d'actine modulent la distribution de certains organites tels que le complexe de Golgi, le réticulum endoplasmique et les mitochondries. De plus, ils participent également au positionnement spatial des RNMS afin qu'ils soient lus par les ribosomes.
L'ensemble du téléphone portable des microfilaments, en particulier ceux qui sont étroitement liés à la membrane plasmique, participent à la formation de membranes cellulaires ondulées des cellules qui ont un mouvement actif constant.
Ils participent également à la formation de microvages et d'autres protubérances communes à la surface de nombreuses cellules.
Exemple de fonctions dans le foie
Les microfilaments participent au processus de sécrétion de bile dans les hépatocytes (cellules hépatiques) et également dans les mouvements péristaltiques (contraction coordonnée) des canaliculi hépatiques.
Ils contribuent à la différenciation des domaines de la membrane plasmique grâce à leur association avec différents éléments cytosoliques et au contrôle qu'ils exercent sur la topographie de ces éléments intracellulaires.
Pathologies connexes
Il existe peu de maladies associées aux défauts primaires dans la structure ou aux protéines et enzymes régulateurs dans la synthèse des microfilaments, bien que ceux-ci soient directement impliqués dans un grand nombre de fonctions.
Le faible indice des maladies et des malformations dans la structure primaire des microfilaments est dû au fait qu'il existe généralement des gènes codants multiples de l'actine et de ses protéines régulatrices, un phénomène appelé "redondance génétique" ".
L'une des pathologies les plus étudiées est la vitrification des ovocytes sur leur cytosquelette, où une interruption est observée dans le réseau de microfilament cortical, ainsi qu'une dépolimérisation et une désorganisation des microtubules de la broche mitotique.
En termes généraux, cette vitrification provoque une dispersion chromosomique, car elle entraîne un désaccord dans le compactage de la chromatine entière.
Les cellules qui ont une plus grande organisation et une proportion de microfilaments dans leur cytosquelette sont des cellules musculaires striées, par conséquent, la plupart des pathologies sont associées à un dysfonctionnement de l'appareil contractile.
Des microfilaments défectueux ou atypiques ont également été associés à une maladie osseuse connue sous le nom de maladie de Paget.
Les références
- Aguilar-Cuenca, R., Llorente-González, C., Vicente, c., & Vicente-manzanares, m. (2017). La dynamique de l'adhésion coordonnée en microfilament entraîne la migration à cellules unicellulaires et façonne les tissus. F1000 RÉSARCHE, 6.
- Deux remèdes, c. g., Chhabra, d., Kekic, m., Doigt, je. V., Tsubakihara, m., Berry, D. POUR., & Nosworthy, n. J. (2003). Protéines de liaison Actine: régulation des microfilaments cytosquelettiques. Revues physiologiques, 83(2), 433-473.
- Guo, h., Fauci, L., Shelley, M., & Kanso, et. (2018). Bistabilité dans la synchronisation des microfilaments actionnés. Journal of Fluid Mechanics, 836, 304-323.
- Lancement., Langer, R., & Vacanti, J. P. (Eds.). (2011). Principes de l'ingénierie tissulaire. Presse universitaire.
- Robbins, J. (2017). Maladies du cytosquelette: les déminopathies. Dans les myopathies cardiosquelettiques chez les enfants et les adultes (pp. 173-192). Presse universitaire.
- « Caractéristiques du grizzli, habitat, nourriture, comportement
- CISTUS CLUSII Caractéristiques, taxonomie, habitat, distribution, utilisations »