

Structure et fonctions de l'hétérochromatine
- 803
- 174
- Raphaël Charles
La hétérochromatine C'est une partie de la chromatine (protéines d'ADN et d'histone) densément emballées de chromosomes eucaryotes. Il est généralement associé aux régions «silencieuses» du génome, c'est-à-dire avec ceux qui sont transcriptionnellement inactifs.
Heitz, en 1928, a été le premier à distinguer deux types différents de chromatine dans les chromosomes eucaryotes pendant l'interface, décrivant l'euchromatine et l'hétérochromatine basée sur son compactage différentiel.
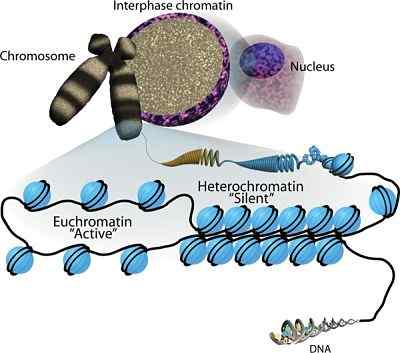
L'organisation de la chromatine dans le noyau (source: sha, k. Et Boyer, L. POUR. La signature de la chromatine des cellules pluripotentes (31 mai 2009), STEMBOOK, ED. La communauté de recherche sur les cellules souches, STEMBOOK, DOI / 10.3824 / STEMbook.1.Quatre cinq.1, http: // www.Carnet de sol.org. [CC par 3.0 (https: // CreativeCommons.Org / licences / par / 3.0)] via Wikimedia Commons) Si les chromosomes eucaryotes sont teints à travers diverses techniques, spécifiques à l'ADN, les observations microscopiques révèlent qu'il existe des régions de ces structures qui sont teintes plus intensément que les autres que les autres. Ces régions correspondent aux régions hypercompactes de l'hectochromatine.
L'hétérochromatinisation de l'ADN, c'est-à-dire que son emballage peut se produire dans une cellule en réponse à différents facteurs et peut être facultatif ou constitutif.
L'hétérochromatine constitutive est une caractéristique permanente qui est généralement héritée, en attendant l'hétérochromatine facultative peut être ou non un chromosome à un moment donné. Le meilleur exemple d'hétérochromatine constitutive est celle de l'un des deux chromosomes X chez les femmes.
Dans les eucaryotes, l'hétérochromatine "stocke" et "compacte" aux grands génomes qui les caractérisent, en particulier les régions qui se composent de séquences répétitives, de fractions résistantes de transposons d'invasion rétro, d'éléments transponables, entre autres.
[TOC]
Structure
L'hétérochromatine n'a pas de structure très différente de celle de la chromatine moins densément emballée, l'euchromatine.
Comprenant cela, il est important de se rappeler que les chromosomes eucaryotes sont composés d'une molécule d'ADN associée à des protéines appelées histones. Huit histones forment un noyau octamérique connu sous le nom de "nucléosome", autour duquel l'ADN est roulé.
Peut vous servir: leucocytes polymorphonucléairesL'association de l'ADN avec les protéines d'histone se produit grâce aux interactions électrostatiques entre les charges positives des déchets de base de ces protéines et les charges négatives des groupes de phosphate de la structure du brin d'ADN.
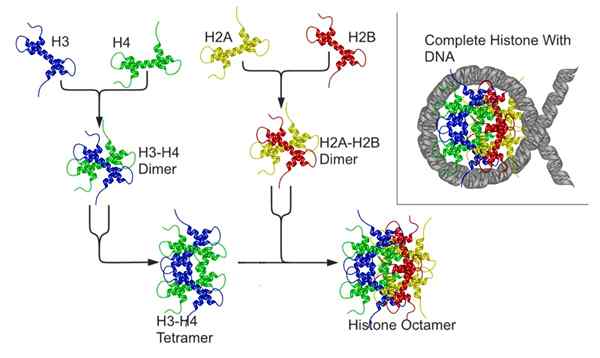 El Nucleosome (Source: Nucleosome_structure.PNG: Richard Wheeler (Zephyris) Travail dérivé (nucléosome-2.Png): rekymanto [cc by-sa 3.0 (https: // CreativeCommons.Org / licences / by-sa / 3.0)] via Wikimedia Commons)
El Nucleosome (Source: Nucleosome_structure.PNG: Richard Wheeler (Zephyris) Travail dérivé (nucléosome-2.Png): rekymanto [cc by-sa 3.0 (https: // CreativeCommons.Org / licences / by-sa / 3.0)] via Wikimedia Commons) - L'octmane Histonas
Chaque octamère d'histone est composé d'un testeur des histones H3 et H4, et de deux jours d'histones H2A et H2B; Autour de chaque noyau d'histone, plus ou moins 146 paires de bases sont logées.
Les nucléosomes s'approchent les uns les autres grâce à la participation d'une autre histone connue sous le nom de Histona de Unión ou Puente (Linker, En anglais), qui est Histon H1.
La chromatine est ensuite composée de nucléosomes successifs qui sont compactés pour former une structure fibreuse d'une épaisseur plus grande mais moins de longueur.
Chaque protéine histone est caractérisée par la présence d'une "queue" d'acides aminés qui peut subir des modifications enzymatiques covalentes. Il a été prouvé que ces modifications affectent le degré d'expression ou le silence des gènes associés aux nucléosomes, ainsi que le niveau de compactage de la chromatine.
En particulier, l'hétérochromatine est caractérisée par l'hypacéthotilation des histones dans toutes les eucaryotes, et par la méthylation de l'histone H3 dans le résidu Lysina 9, uniquement pour les eucaryotes "supérieurs".
Les enzymes responsables de la réalisation de ces modifications sont connues, respectivement, comme les histones Deacel.
En plus des modifications des histones, l'ADN peut également être méthylé, ce qui influence le degré de compactage de la chromatine et correspond à la seconde des deux mécanismes épigénétiques de l'organisation du génome eucaryote.
Peut vous servir: strate basale: caractéristiques et fonctionsOù est l'hétérochromatine?
L'hétérochromatine, comme discuté au début, peut être constitutive ou facultative.
L'hétérochromatine constitutive est particulièrement abondante dans les régions génomiques qui ont une densité élevée de séquences répétitives (telles que les éléments satellites, par exemple), où il y a des éléments transponables abondants, dans les régions centromériques et dans les télomères.
On dit qu'il est constitutif parce que ces régions du génome restent condensées ou compactes pendant la division cellulaire. Dans une cellule qui n'est pas divisée, en revanche, la plupart de l'ADN est euchromatique et il n'y a que des régions bien définies d'hétérochromatine constitutive.
L'hétérochromatine en option est celle trouvée dans les loci qui sont régulées au cours des différentes étapes de développement; Pour ce qu'il représente vraiment, les régions "temporairement condensées" qui peuvent changer en fonction des signaux cellulaires et de l'activité génétique.
Les fonctions
Étant donné que l'hétérochromatine est une partie importante des régions télomériques et centromériques, il exerce des fonctions transcendantales du point de vue de la division cellulaire et de la protection des extrêmes chromosomiques.
Les centromères fonctionnent activement pendant la division cellulaire, permettant le déplacement de chromosomes en double vers les deux pôles de la cellule qui est divisé, tandis que le reste des gènes reste inactif et compact.
Le compactage de régions spécifiques des chromosomes eucaryotes est synonyme de silençage génétique, car le fait que l'hétérochromatine est densément emballée implique l'inaccessibilité des machines transcriptionnelles aux séquences de gènes sous-jacentes.
En ce qui concerne la recombinaison, l'hétérochromatine réprime ce processus, protégeant l'intégrité du génome en interdisant la recombinaison «illégitime» entre les séquences d'ADN répétitives dispersées dans le génome. Ceci est particulièrement important pour le contrôle des éléments transpononables "parasites", qui sont réduits au silence par l'hétérochromatinisation.
Peut vous servir: ProfaseFonctions structurelles
Jusqu'à il y a quelques années, on pensait que l'ADN hétérochromatique était une sorte de "ADN à ordures", car les scientifiques n'ont pas trouvé de fonction spécifique pour les séquences incluses dans ces régions; Rappelons que plus de 80% de l'ADN génomique d'un être humain, par exemple, ne codent pas pour les protéines cellulaires ou pour les molécules d'ARN avec des fonctions de régulation.
Cependant, il est actuellement connu que la formation de l'ADN hétérochromatique facultatif est de la plus haute importance pour la régulation de nombreux processus pendant le développement et la croissance des êtres vivants, et que les régions de l'hétérochromatine constitutive ont un rôle fondamental du point de structure voir.
Il a été suggéré par de nombreux auteurs que l'hétérochromatine peut avoir des fonctions structurelles dans les chromosomes eucaryotes. Cette déclaration est basée sur le fait que les régions hétérochromatiques d'un chromosome donné séparent des parties qui ont des modèles différents de "activité" génétique ".
En d'autres termes, les régions hétérochromatiques servent de «espaceur» entre différentes régions actives transcriptivement, qui peuvent avoir une grande importance du point de vue de la transcription des gènes situés là.
Les références
- Griffiths, un. J., Wessler, s. R., Lewontin, R. C., Gelbart, w. M., Suzuki, D. T., & Miller, J. H. (2005). Une introduction à l'analyse génétique. Macmillan.
- Brun, s. W. (1966). Hétérochromatine. Science, 151 (3709), 417-425.
- Elgin, s. C., & Grewal, s. Toi. (2003). Hétérochromatine: le silence est doré. Biologie actuelle, 13 (23), R895-R898.
- Grewal, s. Toi., & Jia, s. (2007). Révision de l'hétérochromatine. Nature Reviews Genetics, 8 (1), 35.
- Grewal, s. Toi., & Moazed, D. (2003). Hétérochromatine et contrôle épigénétique de l'expression des gènes. Science, 301 (5634), 798-802.
- Hennig, W. (1999). Hétérochromatine. Chromosoma, 108 (1), 1-9.
- « Caractéristiques d'Abelia grandiflora, habitat, culture, soins
- Concept du tropisme, types et exemples »