

Structure des filaments intermédiaires, types, fonctions
- 853
- 192
- Prof Ines Gaillard
Les filaments intermédiaires, Également connu dans la littérature sous le nom de «ifs» (de l'anglais Filames intermédiaires), sont une famille de protéines fibreuses cytosoliques insolubles qui sont présentes dans toutes les cellules eucaryotes multicellulaires.
Ils font partie du cytosquelette, qui est un réseau filamenteux intracellulaire qui est principalement responsable du soutien de la structure cellulaire et de divers processus métaboliques et physiologiques tels que le transport des vésicules, le mouvement et le déplacement cellulaire, etc.
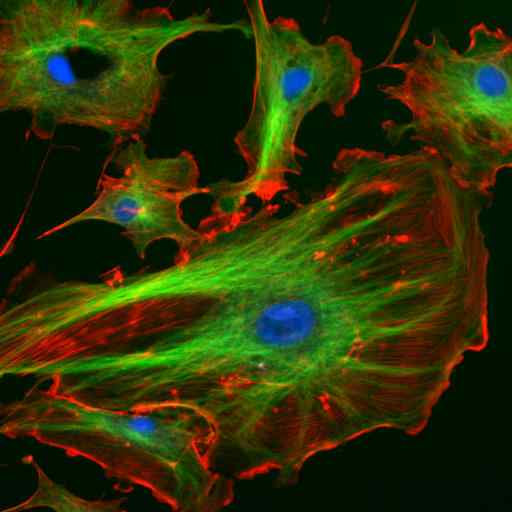
 Microscopie par immunofluorescence de deux protéines des filaments intermédiaires des astrocytes (Vimetina et GFAP) (Source: Gerryshaw [CC BY-SA 3.0 (https: // CreativeCommons.Org / licences / by-sa / 3.0)] via Wikimedia Commons)
Microscopie par immunofluorescence de deux protéines des filaments intermédiaires des astrocytes (Vimetina et GFAP) (Source: Gerryshaw [CC BY-SA 3.0 (https: // CreativeCommons.Org / licences / by-sa / 3.0)] via Wikimedia Commons) Avec les microtubules et les microfilaments, les filaments intermédiaires participent à l'organisation spatiale des organites intracellulaires, dans les processus d'endocytose et d'exocytose et également aux processus de division cellulaire et de communication intercellulaire.
Les premiers filaments intermédiaires qui ont été étudiés et décrits étaient les kératines, l'un des premiers types de protéines dont la structure a été analysée par diffraction des rayons x dans les années 1930.
Le concept de filaments intermédiaires, cependant, a été introduit dans les années 1980 par Lazarides, qui les a décrits comme des «intégrateurs mécaniques de l'espace cellulaire», caractérisé par leur insolubilité et leur capacité à redurer In vitro Après sa dénaturation.
De nombreux auteurs sont considérés comme les éléments de stress "d'amorti" pour les cellules animales, car ce sont des filaments plus flexibles que les microtubules et les microfilaments. Non seulement ils se trouvent dans le cytosquelette, mais ils font également partie du nucléoesqueleto.
Contrairement aux autres composants fibreux du cytosquelette, les filaments intermédiaires ne participent pas directement aux processus de mobilité cellulaire, mais fonctionnent dans le maintien structurel et la résistance mécanique des cellules.
[TOC]
Structure
Source: http: // rsb.Info.NIH.Gov / IJ / Images / [Domaine public] Les filaments intermédiaires ont un diamètre approximatif de 10 nm, caractéristique structurelle par laquelle ils ont été nommés, car leur taille se situe entre les tailles correspondant aux filaments de myosine et d'actine, qui sont environ 25 nm et 7 nm, respectivement.
Ils diffèrent structurellement des deux autres types de filaments de cytosquelette, qui sont des polymères de protéines globulaires, dans lesquels leurs unités constituantes sont des protéines fibreuses α-hélicoïdales de grande longueur qui sont regroupées pour former des structures sous forme de corde ou de corde ou de corde ou de corde.
Toutes les protéines qui constituent les filaments intermédiaires ont une organisation moléculaire similaire, qui se compose d'un domaine α-hélicide ou de «corde» qui a des quantités différentes de segments de «formateurs de bobine» de la même taille.
Ce domaine hélicoïdal est flanqué d'une "tête" non terminale et d'une "queue" non hypoïdale à l'extrémité C-terminale, et les deux varient en taille et en séquence d'acides aminés.
Dans la séquence de ces deux extrêmes, les raisons consensuelles sont communes aux 6 types de filaments intermédiaires connus.
Chez les vertébrés, le domaine «corde» de la protéine des filaments cytosoliques intermédiaires a environ 310 déchets d'acides aminés, tandis que les protéines cytosoliques des invertébrés et le stratifié nucléaire ont environ 350 acides aminés en longueur.
Peut vous servir: Reptiles: quelles sont les caractéristiques, la classification, la reproductionAssemblée
Les filaments intermédiaires sont des structures "auto-assemblage" qui n'ont pas d'activité enzymatique, qui les différencie également de leurs homologues cytosquelettiques (microtubules et microfilaments).
Ces structures sont initialement assemblées sous forme de tétramères de protéines filamenteuses qui les composent sous l'influence de cations monovalentes.
Ces tétramères mesurent 62 nm de long et leurs monomères sont associés les uns aux autres pour former les "unités de longueur" (UFL, de l'anglais Filments d'unité-llegth), Ce que l'on appelle la phase 1 de l'assemblage, qui se produit très rapidement.
Les UFL sont les précurseurs des longs filaments et, puisque les dimères qui les composent se lient dans un antiparalle et par voie échelonnée, ces unités ont un domaine central avec deux domaines flanquants à travers lesquels la phase 2 de l'allongement se produit, où l'union longitudinale D'autres UFL sont donnés.
Pendant ce qui a été appelé comme phase 3 de l'assemblage, le compactage radial du diamètre des filaments se produit, ce qui produit les filaments intermédiaires matures de plus ou moins de 10 nm de diamètre.
Les fonctions
Les fonctions des filaments intermédiaires dépendent considérablement du type de cellule considéré et, dans le cas des animaux (y compris les humains), leur expression est régulée de manière spécifique au tissu, donc elle dépend également du type de tissu que sous étude.
Les épithéliums, les muscles, le mésenchymateux et les glies et les neurones ont différents types de filaments, spécialisés selon la fonction des cellules auxquelles elles appartiennent.
Parmi ces fonctions, les plus importantes sont le maintien structurel des cellules et la résistance à différentes contraintes mécaniques, car ces structures ont une certaine élasticité qui leur permet d'ouvrir différents types de forces imposées aux cellules.
Types de filaments intermédiaires
Les protéines qui constituent des filaments intermédiaires appartiennent à une grande famille hétérogène de protéines filamenteuses qui sont chimiquement différentes mais qui se distinguent en six classes selon leur homologie des séquences (I, II, III, IV, V et VI).
Bien qu'il ne soit pas très courant, différents types de cellules, dans des conditions très particulières (développement, transformation cellulaire, croissance, etc.) Ils peuvent co-exprimer plus d'une classe de protéines formant des filaments intermédiaires
Classe I et II des filaments intermédiaires: acides et kératines de base
Les kératines représentent la plupart des protéines des filaments intermédiaires et, chez l'homme, ceux-ci représentent plus des trois quarts des filaments intermédiaires.
Ils ont des poids moléculaires qui varient entre 40 et 70 kDa et diffèrent des autres protéines de filaments intermédiaires en raison de leur teneur élevée en glycine et en sérine.
Peut vous servir: hérissons: caractéristiques, habitat, reproduction, nourritureIls sont connus sous le nom d'acide et de kératines de base pour leurs points isoélectriques, qui se trouvent entre 4.9 et 5.4 pour les kératines acides et entre 6.1 et 7.8 pour les bases.
Dans ces deux classes, une trentaine de protéines ont été décrites et sont présentes surtout dans les cellules épithéliales, où les deux types de protéines «co-polimérisent» et forment des filaments composés.
De nombreuses kératines de cas de filaments intermédiaires se trouvent dans des structures telles que les cheveux, les ongles, les cornes, les pointes et les griffes, tandis que la classe II est la plus abondante du cytosol.
Classe III des filaments intermédiaires: protéines de type Demine / Vimentin
Les chutes sont une protéine acide de 53 kDa qui, selon son degré de phosphorylation, a des variantes différentes.
Certains auteurs ont également appelé les filaments de la chute sous forme de "filaments intermédiaires musculaires", car leur présence est assez restreinte, bien qu'en petites quantités, tous les types de cellules musculaires.
Dans les myofibrilles, les chutes se trouvent à la ligne Z, il est donc pensé que cette protéine contribue aux fonctions contractiles des fibres musculaires lorsque vous travaillez dans l'union des myofibrillas et de la membrane plasmique.
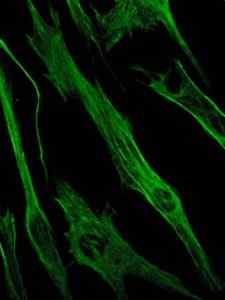 Photographie de la coloration de la protéine de vimentine, une protéine des filaments intermédiaires des cellules épithéliales et embryonnaires (Source: Viktoriia kosach [CC BY-SA 4.0 (https: // CreativeCommons.Org / licences / by-sa / 4.0)] via Wikimedia Commons)
Photographie de la coloration de la protéine de vimentine, une protéine des filaments intermédiaires des cellules épithéliales et embryonnaires (Source: Viktoriia kosach [CC BY-SA 4.0 (https: // CreativeCommons.Org / licences / by-sa / 4.0)] via Wikimedia Commons) À son tour, la vimentine est une protéine présente dans les cellules mésenchymateuses. Les filaments intermédiaires formés par cette protéine sont flexibles et il a été déterminé qu'ils résistent à de nombreux changements de conformation qui se produisent pendant le cycle cellulaire.
Il se trouve dans les fibroblastes, dans les cellules musculaires lisses, dans les globules blancs et d'autres cellules du système circulatoire animal.
Classe IV des filaments intermédiaires: protéines neurofilamentes
Également connue sous le nom de «neurofilaments», ce type de filaments intermédiaires comprend l'un des éléments structurels fondamentaux des axones et des dendrites neuronales; Ils sont souvent associés aux microtubules qui composent également ces structures.
Les neurofilaments d'animaux de vertébrés ont été isolés, déterminant qu'il s'agit d'un triplet protéique de 200, 150 et 68 kDa qui participe à l'assemblage In vitro.
Ils diffèrent des autres filaments intermédiaires dans lesquels ils ont des bras latéraux tels que des «annexes» qui sont projetées de leur périphérie et qui fonctionnent dans l'interaction entre les filaments voisins et d'autres structures.
Les cellules gliales produisent un type spécial de filaments intermédiaires appelés filaments intermédiaires gliaux, qui diffèrent structurellement des neurofilaments, car ils sont composés d'une seule protéine de 51 kDa et qui a différentes propriétés physicochimiques.
Classe V des filaments intermédiaires: filaments de lame nucléaire
Toutes les feuilles qui font partie du Nucleoesqueleto sont en fait des protéines de filament intermédiaires. Ceux-ci ont entre 60 et 75 kDa un poids moléculaire et se trouvent dans les noyaux de toutes les cellules eucaryotes.
Peut vous servir: Trichuris trichiuraIls sont essentiels pour l'organisation interne des régions nucléaires et pour de nombreuses fonctions de cet organelle essentielle pour l'existence des eucaryotes.
Classe VI des filaments intermédiaires: Nestines
Ce type de filaments intermédiaires pèse plus ou moins 200 kDa et est principalement dans les cellules souches du système nerveux central. Ils sont exprimés pendant le développement neuronal.
Pathologies connexes
Il existe de multiples maladies dans les êtres humains qui se rapportent aux filaments intermédiaires.
Dans certains types de cancer tels que les mélanomes malins ou les carcinomes mammaires, par exemple, la co-expression de la vimentine intermédiaire et des filaments de kératine conduit à la différenciation ou à l'interconversion des cellules épithéliales et mésenchymateuses.
Il a été démontré expérimentalement que ce phénomène augmente l'activité migratoire et invasive des cellules cancéreuses, qui a des implications importantes dans les processus métastatiques caractéristiques de cette condition.
Eriksson et les collaborateurs (2009) examinent les différents types de maladies et leur relation avec des mutations spécifiques dans les gènes impliqués dans la formation des six types de filaments intermédiaires.
Les maladies liées aux mutations de gènes codantes pour les deux types de kératine sont l'épidermolyse de Bullast, l'hyperkeratose épidermolytique, la dystrophie cornéenne, les kératodermiers et bien d'autres.
Les filaments intermédiaires de type III sont impliqués dans de nombreuses cardiomyopathies et différentes maladies musculaires liées principalement aux dystrophies. De plus, ils sont également responsables des cataractes dominantes et de certains types de sclérose.
De nombreux syndromes et troubles neurologiques sont liés aux filaments de type IV, tels que Parkinson Parkinson. De même, les défauts génétiques dans les filaments de type V et VI sont responsables du développement de différentes maladies autosomiques et liées au fonctionnement du noyau cellulaire.
Exemple de ceux-ci sont le syndrome de Hutchinson-Gilford Progeria, la dystrophie musculaire Emery-Dreifuss, entre autres.
Les références
- Anderton, B. H. (1981). Filamles intermédiaires: une famille de structures homologues. Journal of Muscle Research and Cell Motility, 2(2), 141-166.
- Eriksson, J. ET., Pallari, h., Robert, D., Eriksson, J. ET., Dechat, t., Sourire, b.,… Goldman, R. D. (2009). Présentation des filamors intermédiaires: de la découverte à la maladie. Le Journal of Clinical Investigation, 119(7), 1763-1771.
- Fuchs, e., & Weber, k. (1994). Filamles intermédiaires: structure, dynamique, fonction et maladie. Anneau. Tour. Biochimie., 63, 345-382.
- Hendrix, m. J. C., Seftor, E. POUR., Chu, et. W., Trevor, k. T., & Seftor, R. ET. B. (mille neuf cent quatre vingt seize). Rôle de la faillament intermédiaire dans la migration, l'invasion et les métastases. Revues de cancer et de métastases, quinze(4), 507-525.
- Herrmann, h., & Aebi, u. (2004). Filames intermédiaires: structure moléculaire, mécanisme d'assemblage et intégration dans des échafaudages fonctionnellement intracellulaires distincts. Revue annuelle de la biochimie, 73(1), 749-789.
- Herrmann, h., & Aebi, u. (2016). Filamles intermédiaires: structure et assemblage. Cold Spring Harbour Perspectives in Biology, 8, 1-22.
- McLean, je., & Lane, B. (Année mille neuf cents quatre-vingts-quinze). Filamors intermédiaires en désir. Opinion actuelle en biologie cellulaire, 7(1), 118-125.
- Steinert, P., & Roop, D. (1988). Biologie moléculaire et cellulaire des films intermédiaires. Revue annuelle de la biochimie, 57(1), 593-625.
- Steinert, P., Jones, J., & Goldman, R. (1984). Filames intermédiaires. Le Journal of Cell Biology, 99(1), 1-6.
- « Caractéristiques gastrotriques, habitat, reproduction, nourriture
- Caractéristiques de la mosquée douce, taxonomie, habitat, utilisations »