

Emballage d'ADN
- 3761
- 994
- Louna Baron
Schéma d'emballage ADN. Source: Thomas Splettstoesser (www.Scistyle.com), cc par 4.0, Wikimedia Commons Qu'est-ce que l'emballage d'ADN?
Il Emballage d'ADN C'est un terme qui définit le compactage contrôlé de l'ADN à l'intérieur de cellule. L'ADN est une molécule extrêmement longue qui, en outre, interagit toujours avec une grande variété de protéines différentes (nucléoprotéines). Avec eux, la chromatine est formée, qui est la substance qui compose les chromosomes.
Pour le traitement, l'héritage et le contrôle de l'expression des gènes que l'ADN adopte une organisation spatiale particulière. La cellule le réalise strictement à contrôler l'emballage d'ADN à différents niveaux de compactage.
Les virus ont différentes stratégies d'emballage de leurs acides nucléiques. L'un des favoris est la formation de spirales compactes. On pourrait dire que les virus sont des acides nucléiques emballés dans les protéines qui les couvrent, protègent et mobilisent.
Dans les procaryotes, l'ADN est associé à des protéines qui déterminent la formation de liens complexes dans une structure appelée nucléoïde. Le niveau de compactage maximal de l'ADN dans une cellule eucaryotique, en revanche, est le chromosome mitotique ou méiotique.
Structure d'ADN
L'ADN est composé de deux bandes antipaallal (ils courent dans la direction opposée) qui forment une double hélice. Chacun d'eux présente un squelette de liaisons phosphodiéster sur lesquelles les sucres liés aux bases d'azote sont joints.
À l'intérieur de la molécule, les bases d'azote d'une bande forment des ponts d'hydrogène (deux ou trois) avec la bande complémentaire.
Dans une molécule comme celle-ci, les angles de liaison les plus importants montrent une rotation gratuite. Les liaisons de base azote-sucre, le phosphate de sucre et la liaison phosphodiéster sont flexibles.
Cela permet à l'ADN, considéré comme une tige flexible, montre une certaine capacité à se plier et à rouler. Cette flexibilité vous permet d'adopter des structures locales complexes et de former des liens d'interaction courtes, moyennes et longues.
Peut vous servir: quelle est la théorie chromosomique de l'héritage? (Sutton et Morgan)Cette flexibilité explique également comment 2 mètres d'ADN peuvent être maintenus dans chaque cellule diploïde d'un être humain. Dans un gamète (cellule haploïde), ce serait un mètre d'ADN.
La nucléoïde bactérienne
Bien que ce ne soit pas une règle inébranlable, le chromosome bactérien existe comme une seule double bande sur la molécule d'ADN à l'éclat.
La double hélice est plus autour de lui-même (plus de 10 pb par retour) produisant ainsi un certain compactage. Des nœuds locaux sont également générés grâce à des manipulations qui sont contrôlées enzimatiquement.
De plus, il y a des séquences d'ADN qui permettent aux domaines de se former en gros liens. La structure résultant du super -clapse et des liens ordonnés est appelée nucléoïde.
Ceux-ci subissent des changements dynamiques grâce à certaines protéines qui offrent une certaine stabilité structurelle au chromosome compacté. Le degré de compactage dans les bactéries et les arches est si efficace qu'il peut y avoir plus d'un chromosome nucléoïde.
Les procaryotes d'ADN nucléoïde compacts au moins environ 1.000 fois. La structure topologique nucléoïde est une partie fondamentale de la régulation des gènes que le chromosome porte. C'est-à-dire que la structure et la fonction constituent la même unité.
Niveaux de compactage chromosomique eucaryote
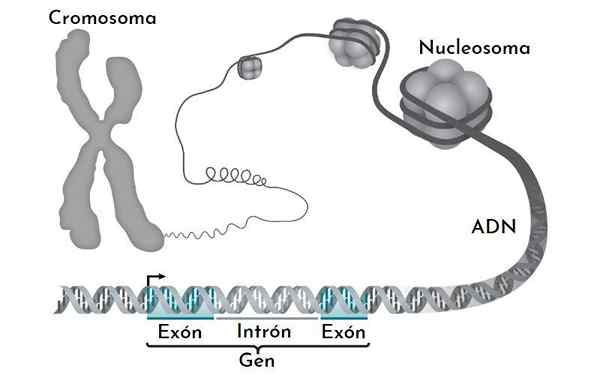
L'ADN dans le noyau eucaryote n'est pas nu. Interagir avec de nombreuses protéines, dont les plus importantes sont les histones. Les histonas sont des petites protéines à chargement positivement qui se lient à l'ADN d'une manière spécifique.
Dans le noyau, ce que nous observons est un ADN complexe: des histones, que nous appelons la chromatine. La chromatine très condensée, qui n'est généralement pas exprimée, est l'hétérochromatine. Au contraire, la chromatine la moins compactée (plus ouverte) ou l'euchromatine est la chromatine avec des gènes qui sont exprimés.
Peut vous servir: ligne pureLa chromatine a plusieurs niveaux de compactage. Le plus élémentaire est le nucléosome. Ils sont suivis par la fibre de solénoïde et les liens de chromatine d'interface. Ce n'est que lorsqu'un chromosome est divisé que les niveaux de compactage maximum sont représentés.
Le nucléosome
Le nucléosome est l'unité de base de l'organisation de la chromatine. Chaque nucléosome est formé par un octa d'histones qui forment une sorte de tambour.
L'octamer est formé par deux copies de chacune des histones H2A, H2B, H3 et H4. Autour d'eux, l'ADN donne presque 1.7 tours. Il est suivi d'une fraction d'ADN libre appelée linker, 20 pb, associée à l'histone H1, puis un autre nucléosome. La quantité d'ADN dans un nucléosome et celle qui le rejoint est d'environ 166 paires de bases.
Cette étape d'emballage de l'ADN compact la molécule environ 7 fois. C'est-à-dire que nous passons un mètre à un peu plus de 14 cm d'ADN.
Cet emballage est possible parce que les histones positives annulent la charge négative de l'ADN et l'auto-réfulsion électrostatique conséquente. L'autre raison est que l'ADN peut être plié de telle manière qu'il peut encercler l'octamère histone.
Fibre de 30 nm
La fibre de comptes dans un collier qui forment de nombreux nucléosomes successifs est en outre inscrit à une structure plus compactée.
Bien que nous ne soyons pas sûrs de ce que la structure adopte vraiment, nous savons qu'elle atteint une épaisseur d'environ 30 nm. Il s'agit de la fibre de 30 nm, et pour sa formation et sa stabilité, l'histone H1 est fondamentale.
La fibre de 30 nm est l'unité structurelle de base de l'hétérochromatine. Celui des nucléosomes laxistes, celui de l'euchromatine.
Liens et virages
La fibre de 30 nm, cependant, n'est pas complètement linéaire. Au contraire, il forme des liens d'environ 300 nm de long, d'une manière sinueuse, sur une matrice protéique peu connue.
Peut vous servir: génétique médico-légale: histoire, objet d'étude, méthodologieCes liens sur une matrice protéique forment une fibre de chromatine plus compacte 250 nm de diamètre. Enfin, ils s'alignent en tant qu'hélice simple de 700 nm d'épaisseur, donnant naissance à l'une des chromatides sœurs d'un chromosome mitotique.
En fin de compte, l'ADN dans la chromatine nucléaire compacte environ 10.000 fois sur le chromosome cellulaire en division. Dans le cœur de l'interface, son compactage est également élevé, car il est environ 1.000 fois par rapport à l'ADN "linéaire".
Le compactage méiotique de l'ADN
Dans le monde de la biologie du développement, il est dit que la gameteogenèse réinitialise l'épigénome. C'est-à-dire qu'il efface les marques d'ADN que la vie de ceux qui ont donné naissance à Gameto ont produit ou expérimenté.
Ces marques incluent la méthylation de l'ADN et les modifications covalentes des histones (code histone). Mais tout l'épigénome n'est pas réinitialisé. Ce qui reste avec les marques sera responsable de l'empreinte génétique paternelle ou maternelle.
La réinitialisation implicite à la gamétoogenèse est plus facile à la voir dans le sperme. Dans le sperme, l'ADN n'est pas emballé avec des histones. Par conséquent, les informations associées à ses modifications dans l'organisme de production, généralement, ne sont pas héritées.
Dans les spermatozoïdes, l'ADN est emballé grâce à l'interaction avec des protéines syndicales non spécifiques à l'ADN, appelées protamines. Ces protéines forment des ponts disulfure entre eux, contribuant ainsi à la formation de couches débordantes qui ne se repoussent pas électrostatiquement.
Les références
- Emballage d'ADN: nucléosomes et chromatine. Récupéré de la nature.com.