

Structure des acides gras, types, fonctions, biosynthèse
- 3119
- 158
- Justine Charpentier
Les des acides gras Ce sont des macromolécules organiques dérivées d'hydrocarbures, qui sont composées de longues chaînes de carbone et d'atomes d'hydrogène qui sont hydrophobes (sont solubles en graisse) et sont la base structurelle des graisses et des lipides.
Ce sont des molécules très diverses qui se distinguent les unes des autres par la longueur de leurs chaînes d'hydrocarbures et la présence, le nombre, la position et / ou la configuration de leurs doubles liaisons.
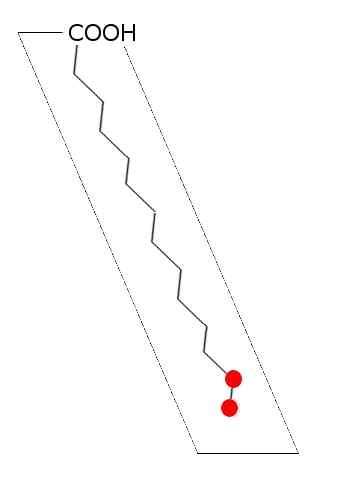
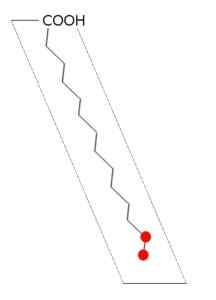 Schéma général d'acide gras saturé (source: Laghi.L [cc by-sa 3.0 (http: // CreativeCommons.Org / licences / by-sa / 3.0 /)] via Wikimedia Commons)
Schéma général d'acide gras saturé (source: Laghi.L [cc by-sa 3.0 (http: // CreativeCommons.Org / licences / by-sa / 3.0 /)] via Wikimedia Commons) Dans les lipides des animaux, des plantes, des champignons et des micro-organismes tels que les bactéries et les levures, plus de 100 types d'acides gras différents ont été décrits et ils sont considérés comme des espèces et des tissus spécifiques dans la plupart des êtres vivants.
Les huiles et les graisses que l'homme consomment quotidiennement sont d'origine animale ou végétale, sont principalement composées d'acides gras.
[TOC]
Introduction
 Le beurre, entre autres, est essentiellement composé d'acides gras (Source: Africa Studio, via Pixabay.com)
Le beurre, entre autres, est essentiellement composé d'acides gras (Source: Africa Studio, via Pixabay.com) Les molécules d'acide gras remplissent des fonctions importantes au niveau cellulaire, ce qui en fait des composants essentiels et, comme certains d'entre eux ne peuvent pas être synthétisés par les animaux, ils doivent les obtenir du régime.
Les acides gras sont rares en tant qu'espèces libres dans le cytosol cellulaire, ils font donc généralement partie d'autres conjugués moléculaires tels que:
- Lipides, dans les membranes biologiques.
- Triglycérides ou esters d'acide gras, qui servent de réserve dans les plantes et les animaux.
- Cixes, qui sont des esters solides d'acides gras à longue chaîne et d'alcools.
- Autres substances similaires.
Chez les animaux, les acides gras sont stockés dans le cytoplasme des cellules comme de petites gouttes de graisse composées d'un complexe appelé triacylglycérol, qui n'est rien de plus qu'une molécule de glycérol à laquelle il a rejoint, dans chacun de ses atomes de carbone, une chaîne d'acide gras par les obligations de type ester.
Alors que les bactéries ont des acides gras courts et généralement monoinsaturés, il est courant.
Structure
Les acides gras sont des molécules amphipatiques, c'est-à-dire qu'elles ont deux régions chimiquement définies: une région polaire hydrophile et une région apolaire hydrophobe.
La région hydrophobe est constituée d'une longue chaîne d'hydrocarbures qui, en termes chimiques, n'est pas très réactive. La région hydrophile, en revanche, est composée d'un terminal du groupe carboxyle (-COH), qui se comporte comme un acide.
Ce groupe carboxyle terminal ou l'acide carboxylique est ionisé en solution, il est hautement réactif (chimiquement parlant) et est très hydrophile, il représente donc un site de jonction covalente entre l'acide gras et d'autres molécules.
La longueur des chaînes d'hydrocarbures d'acides gras a généralement un nombre même d'atomes de carbone, et cela est étroitement lié au processus de biosynthèse par lequel ils sont produits, car leur croissance se produit à partir de paires de carbones.
Les acides gras les plus courants ont des chaînes entre 16 et 18 atomes de carbone et les animaux, ces chaînes ne sont pas ramifiées.
Classification
Les acides gras sont classés en deux grands groupes en fonction de la nature des liens qui les composent, c'est-à-dire, selon la présence de liaisons simples ou de doubles liaisons entre les atomes de carbone de leurs chaînes d'hydrocarbures.
Ainsi, il existe des acides gras saturés et insaturés.
- Les acides gras saturés n'ont que des liaisons en carbone simples et tous leurs atomes de carbone sont "saturés" ou liés aux molécules d'hydrogène.
- Les acides gras insaturés ont une ou plusieurs liaisons en carbone à double carbone et tous ne sont pas attachés à un atome d'hydrogène.
Les acides gras insaturés sont également divisés en fonction du nombre d'insaturation (doubles liaisons) en mononsaturé, ceux avec une seule double liaison, et polyinsaturés, qui en ont plus.
Les acides gras saturés
Ils ont généralement entre 4 et 26 atomes de carbone unis par de simples liens. Son point de fusion est directement proportionnel à la longueur de la chaîne, c'est-à-dire à son poids moléculaire.
Les acides gras qui ont entre 4 et 8 carbones sont liquides à 25 ° C et sont ceux qui composent des huiles comestibles, tandis que ceux avec plus de 10 atomes de carbone sont solides.
Parmi les plus courants, l'acide laurique, qui est abondant dans les graines de palmier et de noix de coco; L'acide palmitique, qui se trouve dans le palmier, le cacao et le beurre de porcs, et l'acide stéarique, qui se trouve dans le cacao et les huiles hydrogénées.
Ce sont des acides gras avec beaucoup plus de stabilité que les acides gras insaturés, en particulier face à l'oxydation, au moins dans des conditions physiologiques.
Merci au fait que les liaisons de carbone simples peuvent tourner gratuitement.
Acides gras insaturés
Ces acides gras sont très réactifs et sujets à la saturation et à l'oxydation. Ils sont communs dans les plantes et les organismes marins. Ceux qui ont une seule double liaison sont connus sous le nom de mononsaturé ou de monoénico-mono-de-forme, tandis que ceux qui en possèdent plus de deux sont appelés polyéniques ou polyinsaturés.
Peut vous servir: ADN polyméraseLa présence de doubles liaisons est courante parmi les atomes de carbone entre les positions 9 et 10, mais cela ne signifie pas que les acides gras monouinsaturés ne sont pas trouvés avec une insaturation dans une autre position.
Contrairement aux acides gras saturés et insaturés ne sont pas répertoriés dans le groupe carboxyle terminal, mais selon la position de la première double liaison C - C. Ainsi, ceux-ci sont divisés en deux groupes, oméga-6 ou ω6 et oméga-3 ou ω3.
Les acides oméga-6 ont la première double liaison dans le carbone numéro 6 et les acides oméga-3 l'ont dans le carbone numéro 3. La dénomination ω est donnée par la double liaison la plus proche du groupe méthyle final.
Les doubles liens peuvent également être trouvés dans deux configurations géométriques appelées "Cis " et "trans".
La plupart des acides gras insaturés naturels ont une configuration "CIS»Et les doubles liaisons d'acides gras présents dans les graisses commerciales (hydrogénées) se trouvent dans "trans".
Dans les acides gras polyinsaturés, deux doubles liaisons sont généralement séparées les unes des autres au moins par un groupe méthyle, c'est-à-dire un atome de carbone attaché à deux atomes d'hydrogène.
Les fonctions
Les acides gras ont de multiples fonctions dans les organismes vivants et, comme mentionné ci-dessus, l'une de leurs fonctions essentielles comme une partie essentielle des lipides, qui sont les principales composantes des membranes biologiques et l'une des trois biomolécules les plus abondantes dans les organismes des organismes vivants en conjonction avec protéines et glucides.
Ce sont également d'excellents substrats d'énergie grâce à laquelle de grandes quantités d'énergie sont obtenues sous forme d'ATP et d'autres métabolites intermédiaires.
Compte tenu du fait que les animaux, par exemple, ne sont pas en mesure de stocker des glucides, les acides gras représentent la principale source de stockage d'énergie qui provient de l'oxydation des sucres en excès.
Les acides gras saturés courtes dans le côlon participent à la stimulation de l'absorption d'eau et des ions de sodium, de chlorure et de bicarbonate; De plus, ils ont des fonctions dans la production de mucus, dans la prolifération des colonocytes (cellules du côlon), etc.
Les acides gras insaturés sont particulièrement abondants dans les huiles comestibles d'origine végétale, qui sont importantes dans l'alimentation de tous les êtres humains.
 Les huiles que nous consommons quotidiennement sont des acides gras (source: Stevepb, via Pixabay.com)
Les huiles que nous consommons quotidiennement sont des acides gras (source: Stevepb, via Pixabay.com) D'autres participent en tant que ligands de certaines protéines avec des activités enzymatiques, ils sont donc importants en ce qui concerne leurs effets sur le métabolisme énergétique des cellules où ils sont trouvés.
Biosynthèse
La dégradation des acides gras est connue sous le nom de β-oxydation et se produit dans les mitochondries des cellules eucaryotes. La biosynthèse, au contraire, se produit dans le cytosol des cellules animales et des chloroplastes (organites photosynthétiques) des cellules végétales.
Il s'agit d'un processus acétyl-CoA, malonyl-CoA et NADPH, il se produit dans tous les organismes vivants et chez les animaux «supérieurs» comme les mammifères. Par exemple, il est très important dans les tissus hépatiques et adipeux, ainsi que dans les glandes mammaires.
Le NADPH utilisé pour cette voie est principalement un produit des réactions d'oxydation dépendantes du NADP de la voie du phosphate de pentose, tandis que l'acétyl-CoA peut provenir de différentes sources, par exemple, de la décarboxylation oxydative du pyruvate, des krebs et β- oxydation des acides gras.
La voie de biosynthèse, comme la β-oxydation, est hautement régulée dans toutes les cellules par des effecteurs alhéstriques et des modifications covalentes des enzymes qui participent au régulation.
-Synthèse malonyl-CoA
L'itinéraire commence par la formation d'un intermédiaire métabolique connu sous le nom de malonyl-CoA d'une molécule acétyl-CoA et est catalysé par une enzyme multifonctionnelle appelée acétyl-CoA carboxilasa.
Cette réaction est une réaction à l'ajout à une molécule carboxyle (-COH, carboxylation) dépendante de la biotine et se produit en deux étapes:
- Tout d'abord, le transfert, dépendant de l'ATP, d'un carboxyle dérivé du bicarbonate (HCO3-) à une molécule de biotine qui se trouve comme un groupe prothétique (non protéine) associé à l'acétyl-coa carboxylase.
- Par la suite, le CO2 est transféré à l'acétyl-CoA et le malonyl-CoA est produit.
-Réactions d'itinéraire
Chez les animaux, la formation de chaînes de glucides d'acides gras se produit davantage par des réactions de condensation séquentielles catalysées par une enzyme multi-critique et multifonctionnelle connue sous le nom d'acide gras synthase.
Esta enzima cataliza la condensación de una unidad de acetil-CoA y múltiples moléculas de malonil-CoA que son producidas a partir de la reacción de la acetil-CoA carboxilasa, proceso durante el cual se libera una molécula de CO2 por cada malonil-CoA que on ajoute.
Les acides gras en croissance sont stérifiés en une protéine appelée "protéine de porteuse acyle" ou ACP, qui forme des thioés avec des groupes acyle. Dans ET. coli Cette protéine est un polypeptide de 10 kDa, mais chez les animaux, il fait partie de la synthase complexe d'acides gras.
Peut vous servir: NoxaLa rupture de ces liaisons Tioéster libère de grandes quantités d'énergie, ce qui permet, thermodynamiquement parlant, la survenue d'étapes de condensation sur la route biosinthétique.
Complexe d'acides gras synthase
Dans les bactéries, l'activité acide synthase correspond en fait à six enzymes indépendantes qui utilisent l'acétyl-CoA et le malonyl-CoA pour former des acides gras et avec lesquels six activités enzymatiques différentes sont associées.
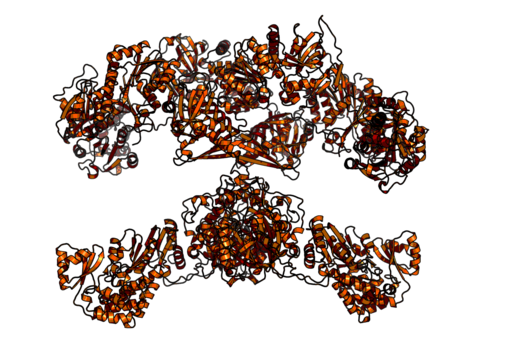 Homodimérique et complexe multifonctionnel Acide gras synthase des animaux (Source: Boehringer Ingelheim [CC BY-SA 4.0 (https: // CreativeCommons.Org / licences / by-sa / 4.0)] via Wikimedia Commons)
Homodimérique et complexe multifonctionnel Acide gras synthase des animaux (Source: Boehringer Ingelheim [CC BY-SA 4.0 (https: // CreativeCommons.Org / licences / by-sa / 4.0)] via Wikimedia Commons) Chez les mammifères, en revanche, l'acide gras synthase est un complexe enzymatique homodimatique multifonctionnel d'environ 500 kDa le poids moléculaire, qui a six activités catalytiques différentes et avec laquelle la protéine porteuse acide est associée.
Étape 1: Réaction du Prowter
Les groupes Tiol dans les déchets de cystéine responsables de l'union des intermédiaires métaboliques à l'enzyme ACP doivent être chargés, avant le début de la synthèse, avec les groupes acides nécessaires.
Pour ce faire, le groupe acétyl-CoA acétyle est transféré au groupe Tiol (-Sh) de l'un des résidus de cystéine de la sous-unité ACP de l'acide gras synthase. Cette réaction est catalysée par la sous-unité ACP-acil-transférase.
Le groupe acétyl est ensuite transféré de l'ACP vers un autre résidu de cystéine dans le site catalytique d'une autre sous-unité enzymatique du complexe connu sous le nom de β-cethoacyl-ACP-Sinta. Ainsi, le complexe enzymatique est "Orge" pour commencer la synthèse.
Étape 2: Transfert des unités malonyl-CoA
Le malonyl-CoA produit par l'acétyl-CoA carboxilasa est transféré au groupe Tiol dans l'ACP et pendant cette réaction, la partie COA est perdue. La réaction est catalysée par la sous-unité malonil-ACP-transférase du complexe de synthase d'acide gras, qui produit ensuite malonil-ACP.
Au cours de ce processus, le groupe malonique est lié à l'ACP et au β-cotoacil-ACP-SITA via un lien d'ester et un autre sulhidl, respectivement.
Étape 3: condensation
L'enzyme β-zoacil-ACP-Suntase catalyse le transfert du groupe acétyle qui l'a rejoint dans le passage de «l'orge» vers le carbone 2 du groupe Malonilo qui, à l'étape précédente, a été transféré à l'ACP.
Au cours de cette réaction, une molécule de CO2 est libérée qui provient du malonyle, qui correspond au CO2 apporté par le bicarbonate dans la réaction carboxylation de la carboxylase acétyl-CoA. Puis, l'acétoacéthyl-ACP se produit.
Étape 4: réduction
La sous-unité β-zoacil-ACP-réductase catalyse la réduction de l'aceoacétyl-ACP dépendant du NADPH.
Étape 5: déshydratation
Cette étape forme le trans-α, le β-acyl-ACP ou le ∆2-insatura-acil-ACP (crrratonil-ACP), produit de la déshydratation du D-β-hydroxybutiril-ACP par action de la sous-unité en colère-ACP -Hydratasa.
Plus tard, le cratonil-ACP est réduit à Buriril-ACP par une réaction dépendante de NADPH catalysée par la sous-unité Angry-AC-réductase. Avec cette réaction, le premier des sept cycles nécessaires pour produire du palmitail-ACP, qui est le précurseur de presque tous les acides gras est terminé.
Comment suivent les réactions de condensation ultérieures?
Le groupe Butirilo est transféré de l'ACP vers le groupe Tiol d'un résidu de cystéine dans le β-cethoacil-ACP-SITA, de sorte que l'ACP est capable d'accepter un autre groupe malonique du malonyl-CoA.
De cette façon, la réaction qui se produit est la condensation de la malonil-ACP avec le butouril-β-cotoacil-ACP-SINTA, qui donne naissance à β-zotohexanoil-ACP + CO2.
Le palmitail-ACP qui provient des étapes suivantes (après l'ajout de 5 autres unités maloniques) peut être libérée en acide palmitique libre grâce à l'activité de l'enzyme thioestérase, il peut être transféré au COA ou incorporer dans l'acide phosphatide pour la voie de synthèse des phospholipides et des triacylglycérides.
 Structure de l'acide palmitique (source: Andel, via Wikimedia Commons)
Structure de l'acide palmitique (source: Andel, via Wikimedia Commons) La synthase d'acide gras de la plupart des organismes s'arrête dans la synthèse du palmitail-ACP, étant donné que le site catalytique de la sous-unité β-cethoacyl-ACP-synthase a une configuration dans laquelle seuls les acides gras de cette longueur peuvent être adaptés.
Comment sont les acides gras du nombre impair d'atomes de carbone?
Ceux-ci sont relativement courants dans les organismes marins et sont également synthétisés par une synthase d'acide gras complexe. Cependant, la réaction «d'orge» se produit avec une molécule plus longue, le propionyl-ACP, de trois atomes de carbone.
Où et comment sont les acides gras des chaînes plus longues?
L'acide palmitique, comme commenté, sert de précurseur pour de nombreux acides gras saturés et insaturés de chaînes plus longues. Le processus "d'allongement" des acides gras se produit dans les mitochondries, tandis que l'introduction de l'insaturation se produit essentiellement dans le réticulum endoplasmique.
De nombreux organismes transforment leurs acides gras saturés en insaturés comme une adaptation à de faibles températures environnementales, car cela leur permet de maintenir le point de fusion des lipides sous la température ambiante.
Propriétés d'acides gras
De nombreuses propriétés des acides gras dépendent de leur longueur de chaîne et de la présence et du nombre d'insaturation:
Peut vous servir: théorie de la synthèse abiotique: caractéristiques principales- Les acides gras insaturés ont des points de fusion plus faibles que les acides gras saturés de la même longueur.
- La longueur des acides gras (le nombre d'atomes de carbone) est inversement proportionnelle à la fluidité ou à la flexibilité de la molécule, c'est-à-dire que les molécules les plus "courtes" sont plus fluides et vice versa.
En général, les substances grasses fluides sont composées d'acides gras avec des chaînes courtes et avec une présence d'insaturation.
Les plantes ont une quantité abondante d'acides gras insaturés, ainsi que des animaux qui vivent à des températures très basses, car ceux-ci, en tant que composants des lipides présents dans les membranes cellulaires, leur donnent une plus grande fluidité dans ces conditions.
Dans des conditions physiologiques, la présence d'une double liaison dans la chaîne d'hydrocarbures d'une graisse.
La présence de doubles liaisons dans les acides gras associés aux molécules lipidiques a des effets directs sur le degré de "emballage" qu'ils peuvent avoir dans les membranes auxquelles elles appartiennent et avec elles ont également des effets sur les protéines membranaires.
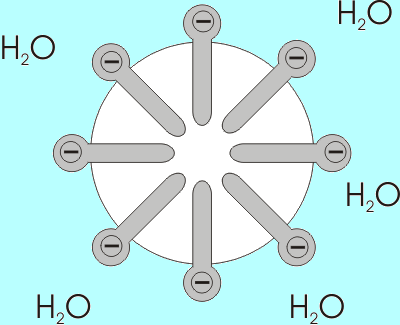 Exemple de la formation d'un myla d'acides gras avec les groupes carboxyliques exposés à l'environnement aqueux (Source: Benutzer: Anderl [CC BY-SA 3.0 (http: // CreativeCommons.Org / licences / by-sa / 3.0 /)] via Wikimedia Commons)
Exemple de la formation d'un myla d'acides gras avec les groupes carboxyliques exposés à l'environnement aqueux (Source: Benutzer: Anderl [CC BY-SA 3.0 (http: // CreativeCommons.Org / licences / by-sa / 3.0 /)] via Wikimedia Commons) La solubilité des acides gras diminue à mesure que la longueur de sa chaîne augmente, ils sont donc inversement proportionnels. Dans les mélanges aqueux et lipidiques, les acides gras sont associés dans des structures appelées micelles.
Une micella est une structure dans laquelle les chaînes aliphatiques des acides gras sont "fermées", "expulsant" toutes les molécules d'eau et à la surface duquel il y a des groupes de carboxyles.
Nomenclature
La nomenclature des acides gras peut être quelque peu complexe, surtout si l'on se réfère aux noms communs qu'ils reçoivent, qui sont souvent liés à une propriété physicochimique, avec l'endroit où ils se trouvent ou d'autres caractéristiques.
De nombreux auteurs considèrent que comme grâce au groupe carboxyle terminal, ces molécules sont ionisées au pH physiologique, il faut les qualifier de "carboxilates" en utilisant la terminaison "Ato ".
Selon le système IUPAC, l'énumération des atomes de carbone d'un gras. La chaîne terminal méthyl contient l'atome de carbone ω.
En général, dans la nomenclature systématique, ils reçoivent le nom de l'hydrocarbure "parental" (hydrocarbure avec le même nombre d'atomes de carbone) et leur résiliation est remplacée "soit" pour "Oico", S'il s'agit d'un acide gras insaturé, la terminaison est ajoutée "Brouillard".
Considérez, par exemple, le cas d'un acide gras de 18 atomes de carbone (C18):
- Étant donné que l'hydrocarbure avec la même quantité d'atomes de carbone est connu sous le nom d'OctaDecano, l'acide saturé est appelé "Acide octadécanoïque" ou bien "Octadécanoato»Et son nom commun est l'acide stéarique.
- Si vous avez une double liaison entre une paire d'atomes de carbone dans sa structure, il est connu sous le nom de "Acide octadécénoïque"
- Si vous avez deux doubles C Res C - C, alors il s'appelle "Acide octadécadiénoïque" Et si vous en avez trois "Acide octadecatrienoique".
Si vous souhaitez résumer la nomenclature, utilisez 18: 0 pour l'acide gras à 18 carbone et aucune double liaison (saturée) et, en fonction du degré d'insaturation, alors au lieu de zéro est écrit 18: 1 pour une molécule avec une molécule avec une insaturation, 18: 2 pour une avec deux insaturation et ainsi de suite.
Si vous souhaitez spécifier entre les atomes de carbone, les doubles liaisons dans les acides gras insaturés, le symbole ∆ est utilisé avec une étude numérique qui indique le lieu de l'insaturation et du préfixe "CIS" soit "trans", en fonction de la configuration de ce.
Les références
- Badui, s. (2006). chimie alimentaire. (ET. Quintanar, Ed.) (4e ed.). Mexique D.F.: Pearson Education.
- Garrett, R., & Grisham, C. (2010). Biochimie (4e ed.). Boston, États-Unis: Brooks / Cole. Cengage Learning.
- Mathews, C., Van Holde, K., & Ahern, k. (2000). Biochimie (3e érigé.). San Francisco, Californie: Pearson.
- Murray, R., Bender, D., Botham, K., Kennelly, P., Rodwell, V., & Weil, P. (2009). Biochimie illustrée de Harper (28e Ed.). McGraw-Hill Medical.
- Nelson, D. L., & Cox, M. M. (2009). Principes de lehninger de la biochimie. Éditions Omega (5e Ed.).
- Rawn, J. D. (1998). Biochimie. Burlington, Massachusetts: Neil Patterson Publishers.
- Tvrzicka, e., Kremmyda, L., Stankova, b., & Zak, à. (2011). Les acides gras comme biocomposs: leur rôle dans le métabolisme humain, la santé et la maladie - une revue. Partie 1: Classification, sources alimentaires et fonctions biologiques. Biomed Pap Med Fac Univ Palacky Olomouc Tchèque Répub, 155(2), 117-130.
- « Structure des glycoprotéines, fonctions, classes et exemples
- L'impact de l'appartenance à la citoyenneté numérique »