

Loi Hardy-Weinberg
- 3949
- 477
- Paul Dumas
La loi de Hardy-Weinberg établit qu'il n'y a pas de variation génétique si la sélection naturelle ou d'autres facteurs n'agissent pas. Source: Wikimedia Commons Qu'est-ce que la loi de Hardy-Weinberg?
La le droit de Hardy-weinberg, Également appelé principe ou équilibre Hardy-Weinberg, il se compose d'un théorème mathématique qui décrit une population diploïde hypothétique avec une reproduction sexuelle qui n'évolue pas, c'est-à-dire que les fréquences alléliques ne changent pas de génération en génération en génération.
Ce principe suppose cinq conditions nécessaires pour que la population reste constante: l'absence de flux de gènes, l'absence de mutations, l'accouplement aléatoire, l'absence de sélection naturelle et une taille de population infiniment grande. En l'absence de ces forces, la population reste en équilibre.
Lorsque l'une des hypothèses ci-dessus n'est pas réalisée, un changement se produit. Pour cette raison, la sélection naturelle, la mutation, les migrations et la dérive génétique sont les quatre mécanismes évolutifs.
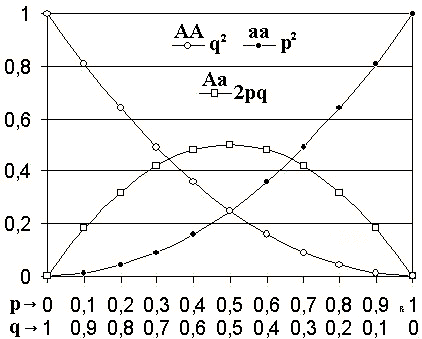
Selon ce modèle, lorsque les fréquences alléliques d'une population sont p et q, Les fréquences génotypiques seront p2, 2pq et q2.
L'équilibre Hardy-Weinberg peut être appliqué dans le calcul des fréquences de certains allèles d'intérêt, par exemple, pour calculer la proportion d'hétérozygotes dans une population humaine.
Nous pouvons également vérifier si une population est en équilibre et proposer une hypothèse de quelles forces agissent dans cette population.
Qu'est-ce que la théorie de Hardy-Weinberg?
La théorie ou l'équilibre de Hardy-Weinberg est un modèle nul qui nous permet de spécifier le comportement des fréquences gènes et alléliques tout au long des générations.
En d'autres termes, c'est le modèle qui décrit le comportement des gènes dans les populations, dans une série de conditions spécifiques.
Notation
Dans le théorème de Hardy-Weinberg, la fréquence allélique de POUR (l'allèle dominant) est représenté par la lettre p, tandis que la fréquence allélique de pour (l'allèle récessif) est représenté par la lettre q.
Les fréquences génotypiques attendues sont p2, 2pq et q2, Pour l'homozygotus dominant (AA), hétérozygote (AA) et homozygotes récessives (AA), respectivement.
Peut vous servir: Holoenzyme: caractéristiques, fonctions et exemplesS'il n'y a que deux allèles dans ledit locus, la somme des fréquences des deux allèles doit nécessairement être égale à 1 (P + Q = 1).
L'expansion du binôme (P + Q)2 Ils représentent les fréquences génotypiques p2 + 2Pq + q2 = 1.
Exemple
Dans une population, les individus qui l'intégrent se croisent pour donner naissance à la progéniture. En général, nous pouvons souligner les aspects les plus importants de ce cycle de reproduction: la production de gamètes, la fusion de ceux-ci pour donner naissance à un zygote et le développement de l'embryon pour donner naissance à la nouvelle génération.
Imaginez que nous pouvons suivre le processus des gènes mendéliens dans les événements susmentionnés. Nous le faisons parce que nous voulons savoir si un allèle ou un génotype augmentera ou diminuera sa fréquence et pourquoi il le fait.
Pour comprendre comment les fréquences gènes et alléliques varient dans une population, nous suivrons la production de gamètes à partir d'un ensemble de souris.
Dans notre exemple hypothétique, l'accouplement se produit au hasard, où tous les spermatozoïdes et ovules sont mélangés au hasard.
Dans le cas des souris, cette hypothèse n'est pas vraie et n'est qu'une simplification pour faciliter les calculs. Cependant, dans certains groupes d'animaux, tels que certains échinodermes et autres organismes aquatiques, les gamètes sont expulsés et entrent au hasard.
Première génération de souris
Maintenant, concentrons notre attention sur un locus spécifique, avec deux allèles: POUR et pour. Suite à la loi énoncée par Gregor Mendel, chaque gamète reçoit un allèle de locus A. Supposons que 60% des ovules et des spermatozoïdes reçoivent l'allèle POUR, Alors que les 40% restants ont reçu l'allèle pour.
Par conséquent, la fréquence des allèles POUR est 0,6 et l'allèle pour C'est 0,4. Ce groupe de gamètes sera trouvé au hasard pour donner naissance à un zygote, ce qui est susceptible de former chacun des trois génotypes possibles? Pour ce faire, nous devons multiplier les probabilités comme suit:
Il peut vous servir: Quintana Roo Flora and FaunaGénotype AA: 0,6 x 0,6 = 0,36.
Génotype AA: 0,6 x 0,4 = 0,24. Dans le cas d'Hétérozygotus, il y a deux formes dans lesquelles il peut être originaire. Le premier, que le sperme transporte l'allèle POUR Et l'ovule l'allèle pour, ou le cas inversé, le sperme pour Et l'ovule POUR. Par conséquent, nous ajoutons 0,24 + 0,24 = 0,48.
Génotype AA: 0,4 x 0,4 = 0,16.
Deuxième génération de souris
Maintenant, imaginons que ces zygotes se développent et deviennent des souris adultes qui produiront à nouveau des gamètes, nous attendrions-nous à nous attendre à ce que les fréquences alléliques soient égales ou différentes de la génération précédente?
Le génotype AA Il produira 36% des gamètes, tandis que les hétérozygotes produiront 48% des gamètes et le génotype AA 16%.
Pour calculer la nouvelle fréquence des allèles, nous ajoutons la fréquence de l'homozygotus plus la moitié des hétérozygotes, comme suit:
Fréquence des allèles POUR: 0,36 + ½ (0,48) = 0,6.
Fréquence des allèles pour: 0,16 + ½ (0,48) = 0,4.
Si nous les comparons aux fréquences initiales, nous nous rendrons compte qu'ils sont identiques. Par conséquent, selon le concept d'évolution, car il n'y a pas de changements dans les fréquences alléliques tout au long des générations, la population est en équilibre, n'évolue pas.
Hardy-weinberg Balance Hypothèses
Quelles conditions la population précédente doit-elle se réunir pour que leurs fréquences alléliques restent constantes avec le passage des générations? Dans le modèle Hardy-Weinberg Balance, la population qui n'évolue pas répond aux hypothèses suivantes:
La population est infiniment grande
- La population doit avoir une taille extrêmement grande pour éviter les effets stochastiques ou aléatoires de la dérive des gènes.
- Lorsque les populations sont faibles, l'effet de la dérive des gènes (changements aléatoires dans les fréquences alléliques, d'une génération à l'autre) en raison de l'erreur d'échantillonnage est beaucoup plus grande et peut produire la fixation ou la perte de certains allèles.
Peut vous servir: fertilisation externeIl n'y a pas de flux de gènes
- Les migrations n'existent pas dans la population, elles ne peuvent donc pas atteindre ou laisser des allèles qui peuvent modifier les fréquences des gènes.
Il n'y a pas de mutations
- Les mutations sont des changements dans la séquence d'ADN et peuvent avoir des causes différentes. Ces changements aléatoires modifient la collection de gènes dans la population, par l'introduction ou l'élimination des gènes dans les chromosomes.
Accouplement aléatoire
- Le mélange des gamètes doit être effectué au hasard - comme l'hypothèse que nous utilisons dans l'exemple des souris-. Par conséquent, il ne devrait pas y avoir de choix de couple parmi les individus de la population, y compris l'endogamie (reproduction des individus liés).
- Lorsque l'accouplement n'est pas aléatoire, il ne provoque pas de changement dans les fréquences alléliques d'une génération à la suivante, mais elle peut générer des écarts par rapport aux fréquences génotypiques attendues.
Il n'y a pas de sélection
- Il n'y a pas de succès reproductif différentiel des individus avec différents génotypes qui peuvent modifier les fréquences des allèles au sein de la population.
En d'autres termes, dans la population hypothétique, tous les génotypes ont la même probabilité de reproduction et de survie.
Lorsqu'une population ne remplit pas ces cinq conditions, le résultat est l'évolution. Logiquement, les populations naturelles ne respectent pas ces hypothèses. Par conséquent, le modèle Hardy-Weinberg est utilisé comme une hypothèse nulle qui nous permet de faire des estimations approximatives des gènes et des fréquences alléliques.
En plus de l'absence de ces cinq conditions, il existe d'autres causes possibles pour lesquelles la population n'est pas en équilibre.
L'un d'eux se produit quand loci sont liés aux phénomènes sexuels ou de distorsion en ségrégation ou Drive méiotique (Lorsque chaque copie d'un gène ou chromosome n'est pas transmise avec une probabilité égale à la génération suivante).
Les références
- Andrews, C. (2010). Le principe Hardy-Weinberg. Connaissances en éducation de la nature.
- Futuyma, D. J. (2005). Évolution. Péchoir.
- Soler, m. (2002). Évolution: la base de la biologie. Projet sud.