

Phase sombre de la photosynthèse
- 1916
- 507
- Paul Dumas
La phase sombre de la photosynthèse est le processus biochimique par lequel les plantes convertissent le dioxyde de carbone en glucose Quelle est la phase sombre de la photosynthèse?
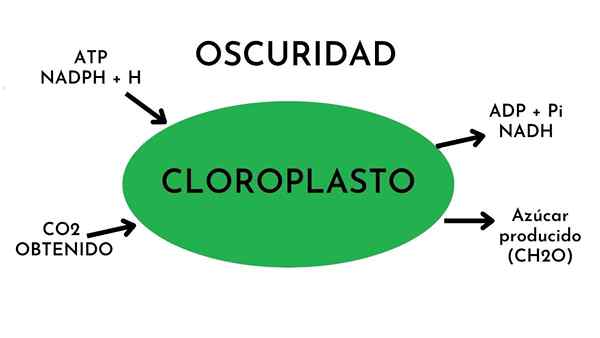
La Phase sombre de la photosynthèse C'est le processus biochimique par lequel les plantes convertissent le dioxyde de carbone en glucose. On dit que la lumière n'est pas nécessaire pour le processus. Il est également connu sous le nom de phase de fixation en carbone ou cycle Calvin-Benson. Ce processus se produit dans le Stroma chloroplaste.
Dans la phase sombre, l'énergie chimique est fournie par des produits générés dans la phase lumineuse. Ces produits sont les molécules d'énergie ATP (adénosine trphosphate) et le NADPH (un porte-électrons réduit).
La matière première fondamentale du processus en phase sombre est le carbone, qui est obtenu à partir de dioxyde de carbone. Le produit final est les glucides ou les sucres simples.
Ces composés carbone obtenus sont la base fondamentale des structures organiques des êtres vivants.
Caractéristiques de la phase sombre de la photosynthèse
- Il est appelé sombre en ne nécessitant pas de participation directe de la lumière du soleil pour son développement. Ce cycle se produit pendant la journée ou la nuit.
- La phase sombre se développe principalement dans le stroma du chloroplaste dans la plupart des organismes photosynthétiques. Le stroma est la matrice qui remplit la cavité interne du chloroplaste autour du système Tilacoïde (où la phase lumineuse est réalisée).
- Dans le stroma se trouvent les enzymes nécessaires pour que la phase sombre se produise. La plus importante de ces enzymes est le Rubisco (biphosphate carboxylase / oxygénase Ribulle.
Mécanismes
Le carbone requis pour le processus est sous la forme de co₂ (dioxyde de carbone) dans l'environnement. Dans le cas des algues et des cyanobactéries, le Co₂ est dissous dans l'eau environnante. Dans le cas des plantes, Co₂ atteint les cellules photosynthétiques à travers des stomates (cellules épidermiques).
Peut vous servir: AshCycle Calvin-Benson
Ce cycle a plusieurs réactions:
Réaction initiale
Le co₂ Il examine un composé accepteur de cinq carbones (Ribulosa 1.5-biphosphate ou RUBP). Ce processus est catalysé par l'enzyme Rubisco. Le composé résultant est une molécule à six carbone.
Il se casse rapidement et forme deux composés de trois carbones chacun (3-phosphoglycérate ou 3pg).
Deuxième processus
Dans ces réactions, l'énergie fournie par l'ATP de la phase lumineuse est utilisée. Une phosphorylation favorisée par l'énergie de l'ATP et un processus de réduction médiée par le NADPH. Ainsi, le 3-phosphoglycérate est réduit au glycéraldéhyde 3-phosphate (G3P).
Le G3P est un sucre Fospatada à trois carbone, également appelé triosa phosphate. Seule une sixième partie du glycéraldéhyde à 3 phosphates (G3P) est transformée en sucres en tant que produit du cycle.
Ce métabolisme photosynthétique est appelé C3, car le produit de base obtenu est un sucre à trois carbone.
Processus final
Les pièces G3P qui ne sont pas transformées en sucres, sont traitées pour former monophosphate Ribulous (croupe). La croupe est un produit intermédiaire qui se transforme en 1,5 biphosphate (RUBP) Ribulous. De cette façon, l'accepteur de Co₂ est récupéré Et le cycle de Kelvin-Benson se ferme.
Du total RUBP produit dans le cycle sur une feuille typique, un seul tiers devient un amid. Ce polysaccharide est stocké dans le chloroplaste comme source de glucose.
Une autre partie est convertie en saccharose (un disaccharide) et transportée vers d'autres organes de la plante. Par la suite, le saccharose est hydrolysé pour former des monosaccharides (glucose et fructueux).
Autres métabolismes photosynthétiques
Dans des conditions environnementales particulières, le processus photosynthétique des plantes a évolué et est devenu plus efficace. Cela a conduit à l'apparition de différentes routes métaboliques pour obtenir des sucres.
Peut vous servir: Tuja occidental: caractéristiques, habitat, homéopathie, cultureMétabolisme C4
Dans des environnements chauds, les stomues des feuilles sont fermées pendant la journée pour éviter la perte de vapeur d'eau. Par conséquent, la concentration de co₂ dans la feuille diminue par rapport à l'oxygène (ou2). L'enzyme Rubisco a une double affinité de substrat: co₂ et o2.
À de faibles concentrations de co₂ et élevé2, Rubisco catalyse la condensation de O2. Ce processus est appelé photoorerspiration et diminue l'efficacité photosynthétique. Pour contrecarrer la photoorerspiration, certains environnements tropicaux ont développé une anatomie et une physiologie photosynthétiques particulières.
Pendant le métabolisme C4, le carbone est fixé dans les cellules mésophyliques et le cycle Calvin-Benson se produit dans les cellules de la gaine chlorophyllienne. La fixation du CO₂ se produit pendant la nuit. Cela ne se produit pas dans le stroma du chloroplaste, mais dans le cytosol des cellules mésophyliques.
La fixation du CO₂ se produit par une réaction de carboxylation. L'enzyme qui catalyse la réaction est la phosphoenolpyruvate carboxylase (pep-carboxylase), qui n'est pas sensible aux faibles concentrations de co₂ Dans la cellule.
La molécule de co-accepteur est l'acide phosphoenolpirúvic (PEPA). Le produit intermédiaire obtenu est l'acide oxaloacétique ou l'oxalacétate. L'oxalacétate est réduit en malato dans certaines espèces de plantes ou d'aspartate (un acide aminé) dans d'autres.
Par la suite, le mal se déplace vers les cellules de pod photosynthétiques vasculaires. Ici, il est décarboxylé et pyruvate et co₂ se produit.
Le Co₂ entre dans le cycle Calvin-Benson et réagit avec le Rubisco pour former PGA. Pour sa part, le pyruvate revient aux cellules mésophyliques, où il réagit avec l'ATP pour régénérer l'accepteur de dioxyde de carbone.
Métabolisme de la caméra
Le métabolisme acide des Crrasulaceae (CAM) est une autre stratégie pour la fixation du CO₂. Ce mécanisme a évolué indépendamment dans divers groupes de plantes succulentes.
Il peut vous servir: dicotylédoneLes plantes CAM utilisent à la fois la route C3 et C4, comme dans les plantes C4. Mais la séparation des deux métabolismes est temporaire.
Le CO₂ est réglé la nuit par l'activité de la pep-carboxylase dans le cytosol et forme de l'oxalacétate. L'oxalacétate est réduit en malato, qui est stocké dans la vacuolie sous forme d'acide formel.
Par la suite, en présence de la lumière, l'acide malique est récupéré de la vacuole. Il est décarboxylé et le CO₂ est transféré au RUBP du cycle Calvin-Benson à l'intérieur de la même cellule.
Les plantes CAM ont des cellules photosynthétiques avec de grandes vacuoles où l'acide rican est stocké, et les chloroplastes où le CO₂ obtenu à partir de l'acide musical est transformé en glucides.
Produits finaux
À la fin de la phase sombre de la photosynthèse, différents sucres sont produits. Le saccharose est un produit intermédiaire qui est rapidement mobilisé des feuilles à d'autres parties de la plante. Il peut être utilisé directement pour obtenir du glucose.
L'amidon est utilisé comme substance de réserve. Il peut s'accumuler sur la feuille ou être transporté vers d'autres organes, tels que les tiges et les racines. Il est maintenu jusqu'à ce qu'il soit nécessaire dans différentes parties de la plante. Il est stocké dans des plastes spéciaux, appelés amyloplastes.
Les produits obtenus à partir de ce cycle biochimique sont vitaux pour la plante. Le glucose produit est utilisé comme source de carbone pour établir des composés, tels que les acides aminés, les lipides ou les acides nucléiques.
D'un autre côté, le produit des sucres de la phase sombre générée représente la base de la chaîne alimentaire. Ces composés représentent des forfaits d'énergie solaire transformés en énergie chimique, utilisés par tous les organismes vivants.
Les références
- Raven, P.H., R.F. Evert et S.ET. Eichhorn (1999). Biologie des plantes. Wh Freeman et Company Worth Publishers.
- Salomon, E.P., L.R. Berg et D.W. Martin (2001). la biologie. McGraw-Hill Inter-American.