

Protéines G Structure, types et fonctions
- 4625
- 191
- Louna Baron
Le g soit Protéines de liaison aux nucléotides de guanine, Ce sont des protéines associées à la membrane plasmique appartenant à une famille de «coupleurs» de signaux qui ont des fonctions importantes dans de nombreux processus de transduction du signal dans les organismes eucaryotes.
Dans la littérature, les protéines G sont décrites comme des commutateurs moléculaires binaires, car leur activité biologique est déterminée par les changements de leur structure donnés par les espèces nucléotidiques auxquelles elles sont capables de rejoindre: les nucléotides de guanosine (diphosphate (PIB) et typosphate (GTP))).
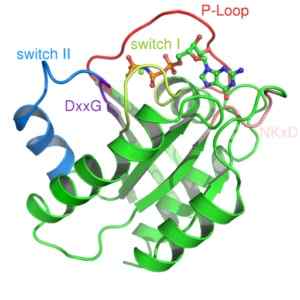
Structure de la protéine RAS, une protéine G monomère (source: Mark 'Absturz' [domaine public] via Wikimedia Commons) Ils sont généralement activés par les récepteurs d'une famille de protéines connues sous le nom de récepteurs couplés GPCR (GPCR, anglais "Récepteurs couplés aux protéines G "), qui reçoivent un signal initial externe et lui font des changements conformationnels qui déclenchent l'activation, qui se traduit par la suite par l'activation d'une autre protéine effective.
Certains auteurs considèrent que les gènes codants de cette famille de protéines ont évolué en duplication et en divergence d'un gène commun ancestral, dont le produit a été affiné et spécialisé de plus en plus.
Parmi la grande variété de fonctions cellulaires que ces protéines ont figurent la translocation de macromolécules pendant la synthèse des protéines, la transduction de signaux hormonaux et des stimuli sensoriels, ainsi que la régulation de la prolifération et de la différenciation cellulaire.
Deux classes ont été décrites à partir de ce type de protéines: petites protéines G et protéines G hétérotrophimériques. La structure à trois premières dimensions d'une protéine G a été obtenue plus d'une décennie d'une petite protéine G connue sous le nom de Ras.
[TOC]
Structure
Structurellement parlant, deux types de g-protéines sont reconnus: petites protéines G et protéines G hétérotrimiques, beaucoup plus complexes.
De petites protéines G sont formées par un seul polypeptide d'environ 200 déchets aminés et environ 20 à 40 kDa, et dans sa structure, il y a un domaine catalytique préservé (le domaine G) composé de cinq hélices α, de six feuilles pliées β et cinq polypeptides boucles.
En revanche, les protéines G hétérotrophimériques sont des protéines membranaires complètes qui sont composées de trois chaînes polypeptidiques, appelées sous-unités α, β et γ.
-La sous-unité α pèse entre 40 et 52 kDa, a une région de liaison aux nucléotides de guanine et a une activité GTPASA pour hydrolyser les liaisons entre les groupes de phosphate GTP.
Les sous-unités α de différentes protéines G partagent certains domaines structurels tels que l'union GTP et l'hydrolyse, mais sont très différents dans les sites de liaison aux récepteurs et les protéines effectrices.
Peut vous servir: broche mitotique-La sous-unité β a un poids moléculaire légèrement inférieur (entre 35 et 36 kDa).
-La sous-unité γ, en revanche, est beaucoup plus petite et a un poids moléculaire approximatif de 8 kDa.
Tous hétérotrophimériques g. Ces deux domaines sont si fortement associés qu'ils sont considérés comme une seule unité fonctionnelle.
Gars
Comme mentionné précédemment, il existe deux types de protéines G: les petits et hétérotriméricas.
Les petites protéines G ont des fonctions de croissance cellulaire, de sécrétion de protéines et d'interaction des vésicules intracellulaires. De leur côté, les protéines G hétérotrophymériques sont associées à la transduction du signal à partir des récepteurs de surface, et agissent également comme des commutateurs qui alternent entre deux états en fonction du nucléotide associé.
Petites protéines G
Ces protéines sont également appelées petites GTPASA, petites protéines de jonction GTP ou superfamille RAS et forment une superfamille indépendante dans la grande classe de GTP hydrolase avec des fonctions régulateurs.
Ces protéines sont très diverses et contrôlent plusieurs processus cellulaires. Ils sont caractérisés par un domaine préservé de liaison au GTP, le domaine "G". L'union de ce phosphate nucléotidique provoque de petits changements conformationnels dans son domaine catalytique dans les protéines GD.
Son activité est intimement liée aux protéines d'activation des GTPASA (GAP, anglais "GTPase activant la protéine ") et avec des facteurs d'échange de nucléotides de Guanina (GEF, anglais "Facteur d'échange de nucléotides de guanine »).
Dans les eucaryotes, cinq classes ou familles de petites protéines G ont été décrites:
-Ras
-Rho
-Rabot
-SAR1 / ARF
-Couru
Les protéines Ras et Rho contrôlent l'expression génétique et les protéines Rho modulent également la réorganisation du cytosquelette. Les protéines du groupe RAB et SAR1 / ARF ont une influence sur le transport vésiculaire et les protéines régulent le transport nucléaire et le cycle cellulaire.
Protéines G hétérotrophimériques
Ce type de protéine mérite également une association avec deux autres facteurs de protéines, de sorte que la voie de signalisation de l'extérieur vers l'intérieur cellulaire se compose de trois éléments dans l'ordre suivant:
- Les récepteurs couplés Une protéines G
- Le g
- Le Protéines ou la Canaux efficaces
Il y a une grande diversité de protéine G hétérotrophimérique et cela est lié à la grande diversité des sous-unités α qui existent dans la nature, dans laquelle seulement 20% de la séquence d'acides aminés est préservée.
Habituellement, les protéines G hétérotrophimériques sont identifiées grâce à la diversité de la sous-unité α, basée principalement sur leurs similitudes fonctionnelles et séquences.
Les sous-unités α constituent quatre familles (la famille GS, la famille GI / O, la famille GQ et la famille G12). Chaque famille est composée d'un «isotype» différent qui, ensemble, ajoute plus de 15 façons différentes des sous-unités α.
Famille Gs
Cette famille contient des représentants qui participent également à la régulation positive des protéines d'adénylate de cyclase et sont exprimées dans la plupart des types de cellules. Il est composé de deux membres: GS et Golf.
L'indice "S " Il se réfère à la stimulation (de l'anglais "Stimulation ") et l'indice "Olf " Il fait référence à "l'odeur" (de l'anglais "Olfaction"). Les protéines de golf sont exprimées en particulier dans les neurones sensoriels responsables de l'odeur.
Famille GE / S
C'est la famille la plus grande et diversifiée. Ils sont exprimés dans de nombreux types de cellules et médient l'inhibition dépendante de la réception de plusieurs types d'adénylciclasses (l'indice "Yo " fait référence à l'inhibition).
Les protéines avec les sous-unités du groupe α sont exprimées en particulier dans les cellules du système nerveux central et ont deux variantes: A et B.
Famille Gq
Les protéines avec cette famille de sous-unité α sont responsables de la régulation de la phospholipase C. Cette famille se compose de quatre membres dont les sous-unités α sont exprimées par différents gènes. Ils sont abondants dans les cellules du foie, dans les cellules rénales et les poumons.
Famille G12
Cette famille s'exprime de manière omniprésente dans les organismes et on ne sait pas avec certitude ce que sont exactement les processus cellulaires régulés par les protéines avec ces sous-unités.
Sous-unités β et γ
Bien que la diversité des structures alpha soit décisive pour l'identification des protéines hétérotimériques, il y a aussi beaucoup de diversité en ce qui concerne les deux autres sous-unités: bêta et gamma.
Les fonctions
Les protéines G participent à la "canalisation" des signaux des récepteurs de la membrane plasmique aux canaux effecteurs ou enzymes.
L'exemple le plus courant de la fonction de ce type de protéine réside dans la régulation de l'enzyme adénylate cyclasa, enzyme responsable de la synthèse de l'adénosine 3 ', 5'-monophosphate ou simplement de l'ampli cyclique, une molécule qui a des fonctions importantes en tant que deuxième messager Dans de nombreux processus cellulaires connus:
Peut vous servir: cellules caliciformes-Phosphorylation sélective des protéines avec des fonctions spécifiques
-Transcription génétique
-Réorganisation du cytosquelette
-Sécrétion
-Dépolarisation membranaire
Ils participent également indirectement à la régulation de la cascade de signalisation des inositols (phosphatidylinitol et de leurs dérivés phosphorylés), qui sont responsables du contrôle des processus dépendants du calcium tels que la chimiotaxie et la sécrétion de facteurs solubles.
De nombreux canaux ioniques et protéines de transport sont directement contrôlés par les protéines G G G. De même, ces protéines sont impliquées dans de nombreux processus sensoriels tels que la vision, l'odeur, entre autres.
Comment travaillent-ils?
Le mode d'interaction d'une protéine G avec des protéines effectrices est typique de chaque classe ou famille de protéines.
Pour les protéines G des récepteurs G-grébranaux (protéines G hétérotrophymériques), l'union d'un nucléotide guanine tel que le PIB ou le diphosphate de guanosín en sous-unité α provoque l'association des trois sous-unités, formant un complexe connu sous le nom de Gαβγ ou G-GDP, qui est lié qui est lié à la membrane.
Si la molécule du PIB est par la suite échangée par une molécule GTP, la sous-unité α attachée au GTP se dissocie des sous-unités β et γ, formant un complexe séparé connu sous le nom de Gα-GTP, qui est capable de modifier l'activité de ses enzymes ou de Diana Conveyor, protéines.
L'activité hydrolytique de cette sous-unité vous permet de terminer l'activation, en échangeant le GTP contre un nouveau PIB, passant à la conformation inactive.
En l'absence des récepteurs excités associés au processus d'échange du PIB du PIB, il est très lent, ce qui signifie que les protéines GD hétérotrimiques n'échangent que le PIB par GTP à une vitesse physiologiquement importante lorsqu'ils sont rejoints à leurs récepteurs excités.
Les références
- Gilman, G. (1987). G protéines: transduiteurs des signaux générés par le récepteur. Revues annuelles en biochimie, 56, 615-649.
- Milligan, G., & Kostenis, et. (2006). Les protéines G hétérotrimiques: une courte histoire. Journal britannique de pharmacologie, 147, 546-555.
- Offermanns, s. (2003). G-protéines comme transduiteurs dans la signalisation transmembranaire. Progrès en biophysique et biologie moléculaire, 83, 101-130.
- Simon, M., Strathmann, m. P., & Gautam, n. (1991). Diversité des protéines G dans la transduction du signal. Science, 252, 802-808.
- Syrovatkina, V., Gai, k. SOIT., Dey, R., & Huang, x. (2016). Régulation, signalisation et fonctions physiologiques des protéines G. Journal of Molecular Biology, 428(19), 3850-3868.
- « Caractéristiques de Crotalus triseriatus, habitat, reproduction
- Caractéristiques du tonnerre, habitat, taxonomie, propriétés »