

Opeón Discovery, modèle, classification, exemples
- 2168
- 577
- Prof Noah Collet
UN Opéron Il se compose d'un groupe de gènes ordonnés consécutivement qui sont régulés les uns avec les autres, qui codent pour des protéines qui sont fonctionnellement liées et qui se trouvent dans tout le génome des bactéries et des génomes "ancestraux".
Ce mécanisme de régulation a été décrit par f. Jacob et J. Monod en 1961, un fait qui leur a valu le prix Nobel de physiologie et de la médecine en 1965. Ces chercheurs ont proposé et démontré le fonctionnement des opérats à travers les gènes qu'ils codent pour les enzymes requises par Escherichia coli Pour l'utilisation du lactose.
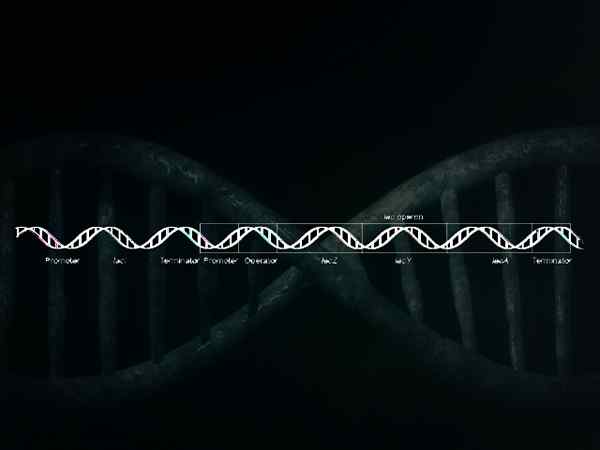
 Schéma graphique d'un brin d'ADN avec les gènes qui comprennent l'opéron lactose (promoteur, opérateur, lacz, lacy, laque et terminateur) (source: llull ~ commonswiki via wikimedia Commons)
Schéma graphique d'un brin d'ADN avec les gènes qui comprennent l'opéron lactose (promoteur, opérateur, lacz, lacy, laque et terminateur) (source: llull ~ commonswiki via wikimedia Commons) Les opérats sont responsables de la coordination de la synthèse des protéines en fonction des besoins de chaque cellule, c'est-à-dire qu'ils s'expriment uniquement pour générer des protéines à l'époque et un endroit exact où ils sont nécessaires.
Les gènes contenus dans les opéras sont généralement. Celles-ci peuvent être la synthèse d'acides aminés, d'énergie sous forme d'ATP, de glucides, etc.
Les opérats se trouvent également souvent dans les organismes eucaryotes, cependant, contrairement aux organismes procaryotes, chez les eucaryotes, la région de l'opérateur n'est pas transcrite en une seule molécule d'ARN messager.
[TOC]
Découverte
La première avancée importante en termes d'exploitation de François Jacob et Jacques Monod était de concentrer le problème de "l'adaptation enzymatique", qui consistait en apparition d'une enzyme spécifique que lorsque la cellule était en présence d'un substrat.
Une telle réponse cellulaire aux substrats a été observée dans les bactéries il y a de nombreuses années. Cependant, les chercheurs se sont demandé comment la cellule a déterminé exactement quelle enzyme devrait synthétiser pour métaboliser ledit substrat.
Jacob et Monod ont observé que les cellules bactériennes, en présence de glucides de type galactose, ont produit 100 fois plus de β-galactosidase que dans des conditions normales. Cette enzyme est ce qui est responsable de la décomposition des β-galactosides afin que la cellule en profite métaboliquement.
Il peut vous servir: Génération filiale: définition et explicationAinsi, les deux chercheurs ont appelé les glucides en galactoside comme "inductances", car ils étaient responsables de l'induction d'une augmentation de la synthèse de la β-galactosidase.
De même, Jacob et Monod ont trouvé une région génétique avec trois gènes qui ont été contrôlés de manière coordonnée: le gène Z, codant pour l'enzyme β-galactosidase; le gène et, codant pour l'enzyme de perméasa lactose (transport de galactosides); et le gène A, qui code pour l'enzyme transacéthylase, qui est également essentielle pour l'assimilation des galactosides.
Grâce à l'analyse génétique ultérieure, Jacob et Monod ont clarifié tous les aspects du contrôle génétique de l'opéron lactose, concluant que le segment des gènes z, et constitue une seule unité génétique avec une expression coordonnée, ce qui était ce qu'ils définissaient comme "opeón".
Modèle d'opéron
Le modèle d'exploitation a été décrit avec précision pour la première fois en 1965 par Jacob et Monod pour expliquer la régulation des gènes transcrits et transcrits pour les enzymes requises en Escherichia coli Afin de métaboliser le lactose comme source d'énergie.
Ces chercheurs ont proposé que les transcriptions du gène ou l'ensemble des gènes qui sont régulés consécutivement par deux éléments: 1) un gène régulateur ou un gène répresseur 2) et un opérateur ou un opérateur de gène opérationnel.
Le gène de fonctionnement est toujours à côté des gènes structurels dont l'expression est responsable de la régulation, tandis que le gène du répresseur code pour une protéine appelée "répresseur" qui se lie à l'opérateur et empêche sa transcription.
La transcription est réprimée lorsque le répresseur est lié au gène de l'opérateur. De cette façon, l'expression génétique des gènes qui codifie les enzymes nécessaires pour assimiler le lactose ne sont pas exprimées et, par conséquent, ne peuvent pas métaboliser ledit disaccharide.
Peut vous servir: patrimoine postmendélien: théories principales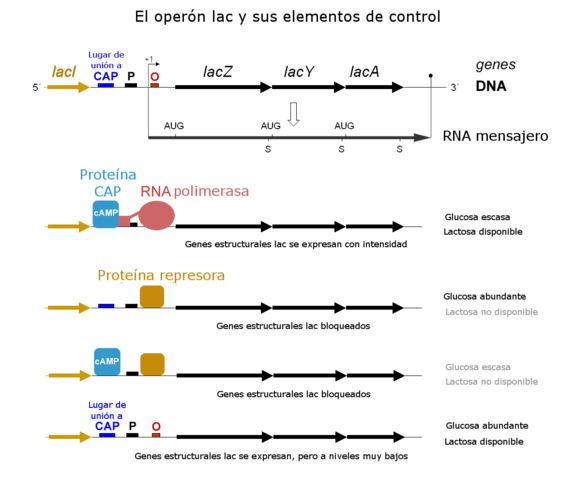 Schéma d'exploitation opérationnel du lactose à travers ses différents éléments de contrôle. Il s'agit de l'opérateur "modèle" utilisé par les professeurs de biologie pour enseigner le fonctionnement de ces gènes (source: Tereseik. Travail dérivé de l'image G3Pro. La traduction espagnole d'Alejandro Porto. [CC par (https: // CreativeCommons.Org / licences / par / 3.0)] via Wikimedia Commons)
Schéma d'exploitation opérationnel du lactose à travers ses différents éléments de contrôle. Il s'agit de l'opérateur "modèle" utilisé par les professeurs de biologie pour enseigner le fonctionnement de ces gènes (source: Tereseik. Travail dérivé de l'image G3Pro. La traduction espagnole d'Alejandro Porto. [CC par (https: // CreativeCommons.Org / licences / par / 3.0)] via Wikimedia Commons) À l'heure actuelle, on sait que la liaison du répresseur à l'opérateur évite, avec les mécanismes stériques, que l'ARN polymérase rejoint le site du promoteur pour commencer à transcrire les gènes.
Le site du promoteur est le "site" qui reconnaît l'ARN polymérase pour rejoindre et transcrire les gènes. Ne pas pouvoir rejoindre ne peut transcrire aucun des gènes de séquence.
Le gène de fonctionnement se situe entre une région génétique de la séquence connue sous le nom de promoteur et de gènes structurels. Cependant, Jacob et Monod n'ont pas identifié cette région en leur temps.
Il est actuellement connu que la séquence complète qui comprend le gène structurel ou les gènes, l'opérateur et le promoteur, est essentiellement ce qui constitue une "opeone".
Classification des opérons
Les opérats ne sont classés que dans trois catégories différentes qui dépendent de la façon dont ils régulent, c'est-à-dire que certains sont exprimés en continu (constitutifs), d'autres ont besoin d'une molécule ou d'un facteur spécifique pour activer (inductible) et d'autres sont exprimés en continu jusqu'à ce que l'inductance soit exprimé (répression).
Les trois types d'opérons sont:
Opeon inductible
Les opeons de ce type sont régulés par des molécules dans l'environnement telles que les acides aminés, les sucres, les métabolites, etc. Ces molécules sont appelées inductances. Si la molécule qui agit comme inducteur, les gènes de l'opéron ne sont pas activement transcrits.
Dans les opérations inductibles, le répresseur libre se lie à l'opérateur et évite la transcription des gènes trouvés dans l'Opeon. Lorsque l'inductance est jointe au répresseur, un complexe est formé qui ne peut pas être joint au répresseur et donc les gènes de l'opéron sont traduits.
Il peut vous servir: fourniture haplonsOpeon répressible
Ces opérons dépendent de molécules spécifiques: acides aminés, sucres, cofacteurs ou facteurs de transcription, entre autres. Ceux-ci sont connus sous le nom de corrigé et agissent totalement en face des inductances.
Ce n'est que lorsque celui corrigé rejoint le répresseur, la transcription s'arrête et donc la transcription des gènes contenus à l'intérieur dans l'opéone ne se produit pas. Ensuite, la transcription d'une opeone répressible ne s'arrête qu'avec la présence du corrigé.
Opeon constitutif
Ces types d'opérates ne sont pas réglementés. Ils sont constamment transcrits activement et, dans un cas de mutation qui affecte la séquence de ces gènes, la durée de vie des cellules qui les contiennent peut être affectée et, généralement, déclenchent la mort cellulaire programmée.
Exemples
Le premier exemple et le plus reconnu de la fonction d'un opéron est l'opérateur LAC (lactose). Ce système est responsable de la transformation du lactose, un disaccharide, dans le glucose et le galactose monosaccharides. Dans ce processus, trois enzymes agissent:
- Β-galactosidase, responsable de la conversion du lactose en glucose et en galactose.
- Permère lactose, chargé de transporter le lactose du milieu extracellulaire dans la cellule et
- Transgéilase, qui appartient au système, mais a une fonction inconnue
L'opéron Trp (tryptophane) de Escherichia coli Contrôle la synthèse du tryptophane, ayant aussi précurseur de l'acide corismique. Dans cette opeone, les gènes de cinq protéines qui sont utilisés pour la production de trois enzymes:
- La première enzyme, codée par les gènes E et D, catalyse les deux premières réactions de la voie du triptophane et est connue sous le nom d'antranilate de synthèse
- La deuxième enzyme est le glycérolphosphate et le catalyz
- La troisième et dernière enzyme est la synthétase tryptophane, en charge de la production du tryptophane à partir du phosphate indol-glycérol et de la sérine (cette enzyme est le produit des gènes B et A) a)
Les références
- Blumenthal, t. (2004). Ouvre dans les eucaryotes. Briefings en génomique fonctionnelle, 3(3), 199-211.
- Gardner, E. J., Simmons, m. J., Snustad, P. D., & Santana Calderón, à. (2000). Principes de la génétique. Principes de la génétique.
- Osbourn, un. ET., & Field, b. (2009). Ouvrir. Sciences de la vie cellulaire et moléculaire, 66 (23), 3755-3775.
- Shapiro, J., Machattie, L., Eron, L., Ihler, g., Ippen, k., & Beckwith, J. (1969). Isolement de l'ADN pur lac opeon. Nature, 224 (5221), 768-774.
- Suzuki, D. T., & Griffiths, un. J. (1976). Une introduction à l'analyse génétique. Wh Freeman et Company.
- « Caractéristiques, fonctions et exemples des cotylédons
- Causes de pénurie d'eau, conséquences, solutions et exemples »